Род Drosera распространен космополитически. Центр разнообразия этого рода находится в Австралии и Новой Зеландии (Juniper, 1989; Баландин, Баландина, 1993). D. rotundifolia является типично арктически-бореальным видом, произрастающим в основном в хвойной зоне Европы и в Сибири (Кац, 1941). D. rotundifolia обычно обитает на верховых болотах, по берегам заболачивающихся озер а также на трясинах - сфагновых болотах на полужидком иле (Masing, 1959; Баранов 1973; Денисова, 1981; Thum, 1986; Redbo-Torstensson, 1994).
В литературе имеются данные о том, что D. rotundifolia обычно растет на самых низких и хорошо увлажненных сфагновых кочках (Crowder et al., 1990, цит. по Redbo-Torstensson, 1994). Однако Thum (1986) отмечает, что на небольшом заиленном болоте в Германии, где автор проводил свои исследования, D. rotundifolia оптимально развивается в наиболее сухих и высоких частях болота. Из этих сведений не ясно, какой тип микрорельефа оптимален для развития D. rotundifolia. Кроме мхов, торфа и ила, D. rotundifolia может произрастать также на влажном песке и "даже в стоячей воде болотных озерков, когда в ней плавают сфагновые мхи" (Masing, 1959). Вероятно, автор имел в виду способность D. rotundifolia произрастать на сплавине, то есть на растительном ковре, лежащем на воде и обычно связанным с берегом (Кац, 1941).
Для D. rotundifolia отмечена положительная связь между количеством пойманных насекомых, размером растения и вероятностью его цветения (Krafft, Handel, 1991; Redbo-Torstensson, 1994).
D. rotundifolia растет обычно в ассоциации со Sphagnum и другими представителями растительных сообществ болота (Masing, 1959; Thum, 1986; Krafft, Handel, 1991). К примеру, в Швеции D. rotundifolia произрастает со S. fuscum, S. rubellum, Eriophorum vaginatum, Andromeda polifolia, Vaccinium oxycoccus (Redbo-Torstensson, 1994).
Межвидовая конкуренция отрицательно влияет на различные морфо-физиологические характеристики насекомоядных растений (Brewer, 1998). Успех ловли насекомых снижается в тех случаях, когда D. rotundifolia находится под пологом растений других видов (Masing, 1959). Особенно тяжело всеми насекомоядными растениями переносится затенение (Chandler, Anderson, 1976; Redbo-Torstensson, 1994; Brewer, 1998). Это объясняется тем, что при недостаточной освещенности снижается интенсивность фотосинтеза, и прекращается образование листовых ловушек (Brewer, 1998). При этом снижается количество пойманных растением насекомых (Masing, 1959). В литературе отсутствуют сведения о влиянии интенсивности освещенности на морфологические характеристики D. rotundifolia. У D. cappilaris в затененных местообитаниях снижается вероятность выживания молодых растений и плотность растений в популяции (Brewer, 1998), а у D. erythrorhyza -- замедляется процесс обновления листьев (Dixon, Pate, 1978).
Межвидовая конкуренция за ресурсы высока в условиях бедных питательными веществами местообитаний, типичных для насекомоядных растений. Это привело к возникновению следующих экологических адаптаций для снижения межвидовой конкуренции. Насекомоядные растения появляются, как правило, на ранних стадиях сукцессии растительных сообществ. Условия обитания насекомоядных растений могут непредсказуемо изменяться под влиянием внешних воздействий (пожары, вырубка леса, покосы, морские волны). При этом роль насекомоядности для растения может меняться в зависимости от содержания в почве питательных веществ и структуры растительных сообществ (Krafft, Handel, 1991).
У D. rotundifolia частота цветения, интенсивность вегетативного роста и успех ловли насекомых зависят от условий обитания (Thum, 1986; Redbo-Torstensson, 1994). Во многих источниках отмечено, что насекомоядные растения приурочены ко влажным местообитаниям (Кац, 1941; Dixon, Pate, 1978; Aldenius et al., 1983; Brewer, 1998).
Насекомоядность рассматривается как приспособление, возникшее в ходе эволюции у растений, обитающих на бедной питательными веществами почве (Masing, 1959; Krafft, Handel, 1991; Brewer, 1998). Насекомоядность является способом для получения дополнительных минеральных веществ, в основном фосфора и азота, а также серы и различных микроэлементов (Chandler and Anderson, 1976; Karlsson and Carlsson, 1984; Redbo-Torstensson, 1994). В характерных для насекомоядных растений олиготрофных местообитаниях большое значение для развития растения имеют минеральные вещества, полученные от пойманных насекомых. При этом эффект от пойманных насекомых проявляется сильнее на более бедных минеральными веществами почвах (Chandler, Anderson, 1976; de Ridder, Dhondt, 1992). Высокое содержание азота в почве приводит к сокращению числа взрослых растений в популяции и положительно влияет на интенсивность вегетативного роста у D. rotundifolia. Однако содержание азота в почве не влияет на успех размножения у этого вида. Количество листьев у D. rotundifolia не зависит от возраста растения и от условий обитания, в том числе от содержания азота в почве (Redbo-Torstensson, 1994).
Из вышесказанного видно, что подавляющее большинство публикаций связано с влиянием насекомоядности на морфологию D. rotundifolia (Krafft, Handel, 1991; Баландин, Баландина, 1993; Redbo-Torstensson, 1994), а также физиологическими и биохимическими аспектами насекомоядности у представителей рода Drosera. Роль насекомоядности в получении D. rotundifolia питательных веществ изучена более, чем сто лет назад, путем лабораторных экспериментов (Kellermann, Raumer, 1878; Darwin, 1878, цит. по собр. соч. Ч. Дарвина, 1908; Busgen, 1883, Порецкий, 1914). Для других видов Drosera в более поздних работах освещены прочие аспекты насекомоядности. К ним относятся видовой состав добычи (Dixon et al. 1980; Thum, 1986), соотношение между потребляемыми растением минеральными веществами (Pate, Dixon, 1978; Chandler, Anderson, 1976) и биохимические аспекты переваривания добычи (Chandler, Anderson, 1976; Chandler, Anderson, 1976b). Некоторые сведения об экологии D. rotundifolia содержаться лишь в работе Thum (1986).
Hanslin и Karlsson (1996) показали, что интенсивность извлечения азота из насекомых у D. rotundifolia в лабораторных условях превышает таковую в естественных условиях более чем в 1,5 раза. Это различие объясняется отсутствием ветра, сильных дождей и резких перепадов температур в лабораторных условиях. Растения, выращенные в лаборатории, отличаются от растений, находящихся в естественных условиях, по характеру метаболизма. Вероятно, это объясняется наличием симбиотической бактериальной флоры у растений, находящихся в естественных условиях (Chandler, Anderson, 1976). Таким образом, проведение исследований в лабораторных условиях не позволяет учитывать ряд факторов, существенно влияющих на жизнедеятельность насекомоядных растений.
В июле и августе 2000 и 2001 годов в рамках экспедиции Гимназии на Юго-Западе исследовали флору 51 острова Керетского архипелага и губы Кив и 39 озер северной части Карельского берега Белого моря. На 31,4% исследованных островов и на берегу 41,0% исследованных озер была обнаружена D. rotundifolia. Отсюда следует, что для района исследований D. rotundifolia является обычным видом, однако распространение этого вида зависит, вероятно, от различных факторов. Целью настоящей работы является изучение влияния факторов внешней среды на различные морфо-физиологические характеристики насекомоядного растения D. rotundifolia.
В период с 23 июля по 11 августа 2001 года в северной части Карельского берега Белого моря провели исследование морфо-физиологических характеристик D. rotundifolia в различных местообитаниях. На обнаруженных популяциях D. rotundifolia случайным образом закладывались площадки размером 20см х 20см (площадью 1/25 кв. м). За период исследований мы однократно обследовали 22 площадки, 7 из которых располагаются на островах губы Кив и Керетского архипелага, а остальные -- на берегах материковых озер. Всего мы исследовали 222 растения. В ходе исследования каждой площадки мы отмечали все виды встреченных на ней растений, включая мхи и лишайники. Для каждого растения измеряли диаметр листовой пластинки наибольшего активного листа, длину его черешка, диаметр листовой розетки, а при наличии цветоноса -- его длину. У D. rotundifolia листовая пластинка имеет округлую форму, ее длина приблизительно равна ее ширине (Кирпичников, 1964; Иконников, 2001). Таким образом, диаметр листовой пластинки полностью характеризует ее размер. Подсчитывали количество пойманных насекомых, а также количество развивающихся и активных листьев на каждом растении. Под развивающимися листьями мы понимаем еще не развернувшиеся листья, не способные ловить насекомых. Активными мы называем зрелые листья, способные ловить насекомых.
Для каждой площадки вычислили процент цветущих растений, процент растений с насекомыми и процент растений, имеющих развивающиеся листья, как отношение числа растений каждого типа к общему числу растений на площадке. Вычислили общую площадь ловчей поверхности растения по формуле S=п(d/2)(d/2)N, где d - диаметр листовой пластинки наибольшего активного листа, N - количество активных листьев.
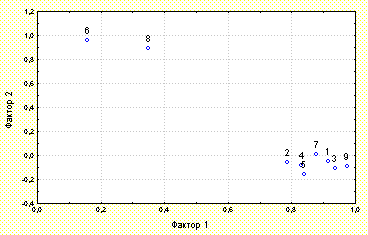
Все характеристики размера D. rotundifolia, к которым относятся количество активных листьев на растении, диаметр листовой пластинки наибольшего активного листа, длина его черешка, а также диаметр листовой розетки, тесно связаны между собой и с общей площадью ловчей поверхности растения (N=222, r=0,53-0,94). Это подтверждается также и факторным анализом усредненных по площадкам данных, при котором все вышеперечисленные характеристики размера растения объединяются в одну группу (рис. 2). Таким образом, площадь ловчей поверхности D. rotundifolia полностью характеризует размер растения. Это позволяет не измерять длину черешка наибольшего активного листа и диаметр листовой розетки при организации дальнейших исследований. Известно, что представители рода Drosera теряют осенью листья и переживают зиму в виде зимующей почки (плотного шарообразного собрания листовых примордий вокруг апикальной меристемы). Каждую весну зимующая почка образует новую листовую розетку (Thum, 1986; Kraft, Handel, 1991). Отсюда следует, что надземная часть D. rotundifolia достигает своего размера за один вегетативный период. Таким образом, общая площадь ловчей поверхности D. rotundifolia является характеристикой интенсивности вегетативного роста этого вида в текущем вегетативном сезоне. Количество развивающихся листьев на растении характеризует интенсивность образования новых листовых ловушек. Количество пойманных D. rotundifolia насекомых является показателем успеха ловли растением насекомых. Вероятность наличия на растении соцветия, а в случае его наличия и длина цветоноса характеризуют интенсивность размножения D. rotundifolia.
Провели кластерный, факторный, дискриминантный и корреляционный анализ полученных в ходе исследований данных (рис. 2). Анализировали параметрические коэффициенты корреляции, поскольку распределение данных было близким к нормальному. Статистическая обработка данных была проведена при помощи пакета STATISTICA (StatSoft, Inc., 1995) в версии STATISTICA 5.0 for Windows.
Проведен факторный анализ значений морфо-физиологических признаков D. rotundifolia, усредненных по модельным площадкам. Выделены два фактора, полностью характеризующих развитие растений. Первый фактор описывает интенсивность размножения и вегетативного роста растений, а также успех ловли растениями насекомых. Второй фактор отражает среднее по площадке количество развивающихся листьев и процент особей на площадке, имеющих развивающиеся листья (рис. 2). Отсюда следует, что количество развивающихся листьев на растении не связано с интенсивностью размножения и вегетативного роста, а также с успехом ловли растением насекомых.
Рис. 2. Результаты факторного анализа значений усредненных по площадкам морфо-физиологических характеристик D. rotundifolia.
Цифрами обозначены морфо-физиологические характеристики D. rotundifolia:
1- диаметр листовой пластинки наибольшего активного листа
2- длина черешка наибольшего активного листа
3- диаметр листовой розетки
4- количество насекомых на растении
5- количество активных листьев на растении
6- количество развивающихся листьев на растении
7- длина цветоноса
8- процент имеющих развивающиеся листья растений на площадке
9- общая площадь ловчей поверхности растения
Площадь ловчей поверхности D. rotundifolia и длина цветоноса у этого вида положительно коррелируют между собой (N=222, r=0,52) и с количеством пойманных растением насекомых (N=222, r=0,61, r=0,48 соответственно). Отсюда следует, что успех ловли растением насекомых, интенсивность его размножения и вегетативного роста у D. rotundifolia связаны между собой. Этот вывод подтверждает данные Krafft и Handel (1991) и Redbo-Torstensson (1994).
При увеличении плотности D. rotundifolia на площадке доля растений с насекомыми на этой площадке уменьшается (N=22, r=-0,50). Это позволяет говорить о наличии внутрипопуляционной конкуренции за насекомых у D. rotundifolia. Плотность растений на площадке отрицательно коррелирует со усредненными по площадке диаметром листовой пластинки наибольшего активного листа и длиной его черешка (N=22, r=-0,50). Таким образом, при увеличении плотности растений длина их листьев уменьшается. При этом предотвращается перекрывание активных листьев, что позволяет снизить конкуренцию за насекомых внутри популяции.
Исследованые нами растения D. rotundifolia в большинстве случаев произрастали на сфагновых сплавинах (36,4% от общего числа исследованных популяций) или болотцах, располагавшихся на сложенных скальными породами возвышенностях (27,3%). Популяции этого вида также были обнаружены на типичных верховых болотах (13,6%), в редкостойном заболоченном сосняке с Betula nana и на каменистой почве (по 9,1%), а также один раз на влажной тропе (4,5%). Таким образом, в большинстве случаев D. rotundifolia произрастает на верховых болотах и в заболоченных местообитаниях, что согласуется с ранее опубликованными данными (Masing, 1959; Баранов 1973; Денисова, 1981; Thum, 1986; Redbo-Torstensson, 1994). Типичным для всех указанных местообитаний является наличие безлесных участков или редкостойного сосняка при отсутствии резких перепадов высот. В таких условиях обитания растения плохо защищены от ветра и не подвержены существенному затенению. Открытость растений ветровым потокам увеличивает вероятность попадания летающих насекомых на листья. Достаточное количество света способствует образованию активных листьев (Masing, 1959; Chandler, Anderson, 1976; Redbo-Torstensson, 1994; Brewer, 1998).
Большинство исследованых площадок располагалось вне сфагновых кочек (63,6%). На болотцах, расположенных на возвышенностях, D. rotundifolia никогда не произрастает на кочках, а на типичных верховых болотах растения всегда находятся на кочках. На сплавинах D. rotundifolia находится и на кочках, и вне кочек. В наблюдаемом распределении D. rotundifolia отсутствует четкая зависимость между расположением растения на кочке или вне ее и условиями местообитания. На основании результатов наших исследований и литературных данных (Thum, 1986; Crowder et al., 1990, цит. по Redbo-Torstensson, 1994) можно предположить, что расположение D. rotundifolia на кочке или вне ее носит случайный характер.
Кластерный анализ морфо-физиологических характеристик всех исследованных растений дал следующие результаты. Все растения разделились на 7 кластеров, в один из которых вошло около одной трети от общего числа исследованных растений. У десяти площадок более 50% растений от общего числа растений на площадке вошло в этот обширный кластер. В подавляющем большинстве случаев растения с одной площадки равномерно распределяются в 2-4 различных кластера. По результатам дискриминантного анализа данных лишь 39,6% D. rotundifolia от общего числа исследованных растений группируются по морфо-физиологическим признакам согласно принадлежности к площадке. Подобные результаты являются аргументом в пользу отсутствия существенного влияния условий обитания на морфо-физиологические характеристики D. rotundifolia.
На большинстве исследованных нами площадок доминировали различные виды Sphagnum (90,9% от общего числа исследованых площадок). В исследованных нами местообитаниях D. rotundifolia обычно произрастает в ассоциации с Vaccinium oxycoccus, Rubus hamaemorus, Empetrum hermaphroditum, Sphagnum fuscum и некоторыми другими видами растений (табл. 1).
Таблица 1. Частота встречаемости различных видов растений (в том числе и лишайников) на исследованных площадках.
| вид | на скольких площадках встречен | процент площадок с этим видом от общего числа площадок (%) |
| Vaccinium oxycoccus | 12 | 54,5 |
| Rubus hamaemorus | 11 | 50 |
| Empetrum hermafroditum | 11 | 50 |
| Sphagnum fuscum | 11 | 50 |
| Andromeda polyfolium | 6 | 27,3 |
| Ledum palustre | 6 | 27,3 |
| вахта | 5 | 22,7 |
| Carex pauciflora | 4 | 18,1 |
| вереск | 4 | 18,1 |
| Carex rariflora | 3 | 13,6 |
| Carex magellanica | 3 | 13,6 |
| Sphagnum rubellum | 3 | 13,6 |
| Pollitrichum communis | 3 | 13,6 |
| сосна | 2 | 9,1 |
| Vaccinium myrtillis | 2 | 9,1 |
| голубика | 2 | 9,1 |
| Betula nana | 2 | 9,1 |
| Sphagnum warnstorfii | 2 | 9,1 |
| Sphagnum fallax | 2 | 9,1 |
| Sphagnum russowii | 2 | 9,1 |
| Molinia caerulea | 1 | 4,5 |
| Cornus suecica | 1 | 4,5 |
| Carex chordorhiza | 1 | 4,5 |
| Carex rostrata | 1 | 4,5 |
| Sphagnum centrale | 1 | 4,5 |
| Sphagnum magellanum | 1 | 4,5 |
| Sphagnum cuspidatum | 1 | 4,5 |
| Pohlia nutans | 1 | 4,5 |
| Cladonia ringoferinum | 1 | 4,5 |
| Ptilidium pulcherinum | 1 | 4,5 |
| Cefallozium sp. | 1 | 4,5 |
| Calypogynum sp. | 1 | 4,5 |
| Microlepidosum sp. | 1 | 4,5 |
Произрастание D. rotundifolia на большинстве исследованных площадок в ассоциации с растениями болотных сообществ согласуется с ранее опубликованными данными (Masing, 1959; Thum, 1986; Krafft, Handel, 1991, Redbo-Torstensson, 1994). Мы не отмечали факта произрастания D. rotundifolia на песке, что не подтверждает данных Masing (1959).
Рядом с модельными площадками, заложенными на берегах озер Лапоть, Эхо, Рибозеро, Верхнее Пулонгское и Левое, произрастал другой вид Drosera, D. anglica Huds. На озерах Эхо, Лапоть и Рибозеро количество развивающихся листьев у D. rotundifolia заметно превышало среднее по всем исследованным площадкам количество листьев этого типа. Среднее по площадке количество развивающихся листьев у D. rotundifolia на озерах Эхо, Лапоть и Рибозеро составляло 0,7-0,8 штук на растение (при среднем по всем площадкам 0,4+/-0,1 штук на растение), процент растений с развивающимися листьями от общего количества растений на площадке варьировал от 60,0 до 77,8% (при среднем по всем площадкам 32,6+/-6,4%). На озерах Верхнее Пулонгское и Левое наблюдали высокие по сравнению с другими площадками показатели интенсивности размножения и вегетативного роста, а также успеха ловли насекомых (рис. 3). Как было показано выше, эти показатели наряду с количеством развивающихся листьев на растении полностью характеризуют развитие D. rotundifolia. Таким образом, растения D. rotundifolia на площадках, рядом с которыми произрастает D. anglica, достоверно отличаются по своим морфо-физиологическим характеристикам от растений с других площадок.
Рис. 3. Усредненные по площадке показатели интенсивности размножения и вегетативного роста, а также успеха ловли насекомых у D. rotundifolia.
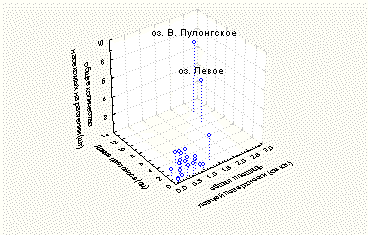
Известно, что в местах совместного произрастания D. rotundifolia и D. anglica встречается гибрид между этими двумя видами, D. х obovata Mert. et Koch. Кроме того, в этом случае образуются переходные формы между D. x obovata и D. rotundifolia, практически не отличимые от D. rotundifolia в полевых условиях (Wood, 1955; Раменская, Андреева, 1982). Возможно, на этих площадках мы исследовали не чистые линии D. rotundifolia, что отразилось на морфо-физиологических характеристиках растений. На площадках на берегу озер Верхнее Пулонгское и Левое отсутствовали обычные для района исследований (то есть встреченные более, чем на 15% исследованных площадок) виды растений, что может свидетельствовать об уникальных условиях обитания на этих площадках. Вероятно, это является еще одной причиной высоких по сравнению с растениями на других площадках показателей интенсивности размножения и вегетативного роста, а также успеха ловли насекомых у D. rotundifolia на озерах Левое и Верхнее Пулонгское.
Диаметр наибольшего активного листа у D. rotundifolia при наличии на площадке Andromeda polifolia больше, чем на тех площадках, где A. polifolia отсутствует (0,4+/-0,03см и 0,3+/-0,01см соответственно). В экспериментах Redbo-Torstensson (1994) наблюдалось увеличение плотности A. polifolia при добавлении в почву азота. Возможно, A. polifolia является индикатором повышенного содержания азота в почве. На основе вышеизложенного можно предположить, что размер D. rotundifolia положительно зависит от содержания азота в почве.
Площадь ловчей поверхности D. rotundifolia при наличии на площадке Carex pauciflora выше, чем на площадках, где C. pauciflora отсутствует (средние значения 1,14+/-0,16 и 0,62+/-0,05 соответственно). Таким образом, C. pauciflora положительно влияет на площадь ловчей поверхности D. rotundifolia. Считается, что C. pauciflora является типичным растением влажных местообитаний (Кац, 1941). Отсюда следует, что площадь ловчей поверхности у D. rotundifolia положительно зависит от степени увлажненности местообитания. Такая зависимость объясняется тем, что при увеличении общей площади поверхности активных листьев потребность D. rotundifolia в воде существенно возрастает. Это происходит из-за того, что активные листья, покрытые секреторными железками, характеризуются высоким уровнем транспирации. Эта гипотеза позволяет обосновать сведения, содержащиеся в работах ряда авторов, о приуроченности насекомоядных растений ко влажным местообитаниям (Кац, 1941; Dixon, Pate, 1978; Aldenius et al., 1983; de Ridder, Dhondt, 1992; Brewer, 1998).
Плотность растений на исследованных площадках колебалась от 75 до 925 растений на кв. м, и в среднем составляла 256+/-43,8 растений на кв. м. По другим данным интервал значений плотности D. rotundifolia не так велик: от 380 до 520 растений на м кв. (Redbo-Torstensson, 1994). Возможно, это объясняется меньшим, чем в настоящей работе, разнообразием исследованных этим автором местообитаний. На небольшом заиленном болоте в Германии средняя плотность D. rotundifolia составляет 405 растений на м кв. (Thum, 1986), что значительно превышает полученное нами значение. Из вышеизложенного видно, что плотность D. rotundifolia значительно варьирует в зависимости от условий обитания.
На площадке, расположенной на о. Плоская Луда, наблюдалась высокая по сравнению с другими площадками плотность растений (700 особей на м кв.). На этой площадке отсутствовали виды рода Sphagnum и произрастали Cornus suecica, лишайник Cladonia ringoferinum и печеночный мох Ptilidium pulcherinum. Подобное растительное сообщество не наблюдалось на других площадках и, вероятно, служит индикатором уникальных условий обитания. Лишайники рода Cladonia считаются типичными растениями очень сухих местообитаний (Кац, 1941). Можно предположить, что плотность D. rotundifolia увеличивается в условиях сухих местообитаний. Однако эта гипотеза требует дальнейшей проверки.
Самая высокая плотность D. rotundifolia (925 особей на м кв.) за счет высокой доли молодых растений (81,1% от общего числа растений на площадке) отмечена на площадке на берегу оз. Плотничное. На этой площадке в отличие от прочих площадок доминирует S. fallax, что, возможно, является индикатором уникальных условий местообитания. Однако нам не удалось выявить внешние факторы, обуславливающие чрезвычайно высокую плотность D. rotundifolia на этой площадке.
1. Успех ловли насекомых, интенсивность размножения и вегетативного роста, а также количество развивающихся листьев у D. rotundifolia полностью характеризуют развитие этого вида.
2. У D. rotundifolia существует внутрипопуляционная конкуренция за насекомых.
3. Обычно D. rotundifolia произрастает на верховых болотах и в заболоченных местообитаниях.
4. Размер D. rotundifolia положительно зависит от содержания азота в почве.
5. Площадь ловчей поверхности у D. rotundifolia положительно зависит от степени увлажненности местообитания.
6. Плотность D. rotundifolia значительно варьирует в зависимости от условий обитания.
7. Не всегда удается выявить внешние факторы, существенно влияющие на морфо-физиологические характеристики D. rotundifolia.
Дарвин Ч. Насекомоядные растения // Собрание сочинений Чарльза Дарвина. Т. 4, Ч. 2. М., 1908. С. 145-154.
Баландин С.А., Баландина Т.П. Росянка круглолистная // Биологическая флора Московской области. Вып. 9, Ч. 2. М., 1993. С. 31-38.
Иконников С.С. Сем. Droseraceae Salisb. -- Росянковые // Флора Восточной Европы. Т. 10. Спб., 2001. С. 302--305.
Кац Н.Я. Болота и торфяники. М., 1941. С. 6-10, 53-61.
Кирпичников М.Э. Сем. Droseraceae -- Росянковые // Маевский П.Ф. Флора средней полосы европейской части СССР. 9-е изд. Л., 1964. С. 355.
Порецкий С.А. Хищные растения. Спб., 1914. С.12-15.
Раменская М.Л., Андреева В.Н. Определитель высших растений Мурманской области и Карелии. Л., 1982. С. 251.
Aldenius J., Carlsson B., Karlsson S. Effects of insect trapping on growth and nutrient content of Pinguicula vulgaris L. in relation to the nutrient content of the substrate // New Phytologist. 1983. Vol. 93. P. 53-59.
Brewer J.S. Effects of competition and litter on a carnivorous plant, Drosera cappilaris (Droseraceae) // American Journal of Botany. 1998. Vol. 85. N 11. P. 1592-1596.
Busgen M. Die Bedeutung des Insektenfangs fur Drosera rotundifolia L. // Botanische Zeitung. 1883. Vol. 41. N 35. P. 569-577.
Chandler G.E., Anderson J. W. Studies on the nutrion and growth of Drosera species with reference to the carnivorous habit // New Phytologist. 1976. Vol. 76. P. 129-141.
Chandler G.E., Anderson J.W. Uptake and metabolism of insect metabolites by leaves and tentacles of Drosera species // New Phytologist. 1976b. Vol. 77. P. 625-634.
De Ridder F., Dhondt A.A. A positive correlation between naturally captured prey, growth and flowering in Drosera intermedia in two contrasting habitats // Belg. Journ. Bot. 1992. Vol. 125. N 1. P. 33-40.
Dixon K.W., Pate J.S. Phenology, morphology and reproductive biology of the tuberous sundew, Drosera erithrorhiza Lindl. // Australian Journal of Botany. 1978. Vol. 26. P. 441-459.
Dixon K. W., Pate J.S., Bailey W.J. Nitrogen nutrition of the tuberous sundew, Drosera erithrorhiza with special reference to catch of arthropod fauna by its glandular leaves // Australian Journal of Botany. 1980. Vol. 28. P. 283-297.
Hanslin H.M., Karlsson P.S. Nitrogen uptake from prey and substrate as affected by prey capture level and plant reproductive status in four carnivorous plant species // Oecologia (Berlin). 1996. Vol. 106. P. 370-375.
Juniper B.E. Carnivorous plants. L., 1989. 353 p.
Karlsson P.S., Carlsson B. Why does Pinguicula vulgaris L. trap insects? // New Phytologist. 1984. Vol. 97. P. 25-30.
Kellerman C., Rauner E. Vegetationsversuche an Drosera rotundifolia mit und ohne Fleischfutterung // Botanische Zeitung. 1878. Vol. 36. N 14. P. 209-218.
Krafft C.C., Handel S.N. The role of carnivory in the growth and reproduction of Drosera filiformis and D. rotundifolia // Bulletin of the Torrey Bot. Club. 1991. Vol. 118. N 1. P. 12-19.
Masing V. Huulheum - Darvin Lemmikmaim // Eesti loodus. 1959. N 6.
Pate J.S., Dixon K.W. Mineral nutrition of Drosera erythrorhiza with special reference to its tuberous habit // Australian Journal of Botany. 1978. Vol. 26. P. 455-464.
Redbo-Torstensson P. The demographic consequences of nitrogen fertilization of a population of sundew, Drosera rotundifolia.// Acta Bot. Neerl. 1994. Vol. 43. N 2. P. 175-188.
Thum M. Segregation of habit and prey in two sympatric carnivorous plant species, Drosera rotundifolia and Drosera intermedia // Oecologia. 1986. Vol. 70. P. 601-605.
Wood C.E. Evidence for the hybrid origin of Drosera anglica // Rhodora. 1955. Vol. 57. P. 105-130.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |