Род Drosera L., принадлежащий к семейству Droseraceae, насчитывает более 90 видов и распространен космополитически. Центр разнообразия этого рода находится в Австралии и Новой Зеландии (Juniper, 1989).
Представители рода Drosera - многолетние, как правило, болотные травянистые растения с коротким ползучим или клубневидным корневищем и прикорневой розеткой черешковых листьев (Денисова, 1981; Баландин, Баландина, 1993). Листья росянок имеют короткие железистые ловчие волоски в центре листовой пластинки и более длинные волоски по ее краям. Головку волоска окружает капля густой липкой слизи. У рода Drosera для выделения слизи и ферментов и для всасывания питательных веществ служит один и тот же тип железок. Мелкие членистоногие (двукрылые, перепончатокрылые, ногохвостки, пауки, клещи и другие) садятся или вползают на лист и прилипают к нему. Все волоски листа изгибаются навстречу добыче и обволакивают ее слизью, в которой содержатся вещества, оказывающие на насекомых парализующее действие, и пищеварительные ферменты. Край листа медленно загибается и закрывает добычу, которая вскоре начинает перевариваться. Когда переваривание закончится, лист раскрывается, волоски выпрямляются, головки их оказываются непокрытыми каплями выделений и непереваренный наружный скелет насекомого удаляется с листа ветром (Дарвин, 1908; Пельцих, 1917).
Установлено, что железистые волоски листа приводятся в движение в тех случаях, когда вслед за механическим раздражением листа следует его химическое раздражение, вызываемое веществами, диффундирующими из тела пойманного насекомого (Холодный, 1948).
Три изучаемых нами вида Drosera отличаются друг от друга в основном формой, размером и положением листовой пластинки. У D. rotundifolia L. листья приближены к земле, при этом черешок резко переходит в листовую пластинку, длина которой несколько меньше ширины. У D. anglica Huds. черешок, расположенный почти перпендикулярно субстрату, постепенно переходит в листовую пластинку, длина которой превышает ширину в 5-10 раз. D. x obovata Mert. et Koch является гибридом D. anglica и D. rotundifolia (Wood, 1955). У этого вида черешок плавно переходит в листовую пластинку, которая несколько приподнята над субстратом. Наибольшая ширина листовой пластинки немного превышает ее длину и расположена в верхней части листа.
Pinguicula vulgaris L. - циркумполярный вид, относящийся к семейству Lentibulariaceae. Жирянка является многолетним травянистым растением с прикорневой розеткой листьев и слабо развитой корневой системой. Этот вид встречается на влажных лугах и болотах. Растение несет в среднем 6-10 довольно толстых, продолговатых, светло-зеленых листьев, почти не имеющих черешка. Молодые листья глубоко вогнуты и торчат вверх, более старые, наружные, плоские или выпуклые и лежат на земле (Дарвин, 1908, Juniper, 1983). Слизевые железки располагаются на верхней стороне листьев, на лепестках, чашелистиках, а также на цветоносе. На 1мм2 в среднем находится 6-8 железок. Функции привлечения добычи и ее переваривания разделены между железками двух разных типов. Слизевые волосковидные железки секретируют ловчую слизь и откладывают ее на ловчие головки. Блестящие капли слизи приманивают насекомых, в то же время, обладая клеящими свойствами, они удерживают пойманную добычу на листовой пластинке, а сидячие пищеварительные железки наиболее активны в пищеварении и всасывании веществ (Муравник, 1988).
Количество пойманных насекомых влияет на вегетативный рост и на вероятность цветения, а также на другие характеристики видов Drosera (Chandler, Anderson, 1976; Karlsson, Carlsson, 1984; Small et al., 1987; Krafft, Handel, 1991; Redbo-Torsensson, 1994; Brewer, 1998). При этом эффект от пойманных насекомых проявляется слабее на более богатой минеральными веществами почве. Надо отметить, что количество пойманных насекомых зависит от погодных условий. Если насекомых мало, то различия в успехе ловли насекомых разными растениями становятся заметнее (De Ridder, Dhondt, 1992). Влияние насекомых на жизнедеятельность зависит и от размера растения. У крупных экземпляров при достаточном количестве насекомых вероятность цветения выше, чем у мелких (De Ridder, Dhondt, 1992; Worley, Harder, 1999).
У Pinguicula vulgaris количество листьев на годичном побеге складывается на ранних стадиях развития и не зависит от количества пойманных насекомых (Karlsson, 1986; Worley, Harder, 1999).
Сравнительный анализ влияния успешности ловли насекомых на морфофизиологические характеристики разных видов насекомоядных растений в литературе отсутствует.
О прочих факторах, влияющих на интенсивность вегетативного роста и размножения у насекомоядных растений, единого мнения не существует. De Ridder и Dhondt (1992) полагают, что на вегетативный рост Drosera intermedia могут влиять условия окружающей среды. По мнению Redbo-Torsensson (1994), количество листьев и цветков у D. rotundifolia не зависит от внешних условий. Успех плодоношения и интенсивность вегетативного роста Pinguicula vulgaris коррелирует с плодородностью почвы по данным Worley и Harder (1999). По другим данным (Aldenius et al., 1983; Molau, 1993), биомасса и интенсивность размножения этого вида зависят и от других характеристик местообитания. При этом важнейшим лимитирующим фактором является доступность воды, так как листья, покрытые секреторными железками, обладают высоким уровнем транспирации (Karlsson, 1986).
Следует отметить, что большинство цитируемых зарубежных работ основаны на исследованиях, производимых над насекомоядными растениями в лабораторных условиях, что значительно суживает область применения сделанных там выводов. В отечественной литературе подобные исследования вообще отсутствуют.
Целью нашего исследования являлось выявление связи между интенсивностью вегетативного роста и размножения и успехом ловли насекомых, а также условиями обитания у различных видов насекомоядных растений. Мы анализировали результаты синхронных ежедневных наблюдений над популяциями насекомоядных растений в полевых условиях.
С 25 по 29 июля 1999 года и с 12 июля по 5 августа 2000 года вели ежедневные наблюдения за насекомоядными растениями, принадлежащими к родам Drosera и Pinguicula в Лоухском районе республики Карелия в окрестностях губы Чупа Белого моря (62 30' с. ш., 32 35' в. д.) (карта).
В 1999 году пометили 23 растения из популяции Drosera rotundifolia, расположенной на мысе Иванов Наволок. Данная популяция произрастала на олиготрофном болотце, располагавшимся на сложенной базальтовыми породами возвышенности. Вероятность того, что метки влияли на успех ловли насекомых, мала, так как метки были очень легкими и прикреплялись на цветоносе. В течение пяти дней вели наблюдение за поведением всех меченых растений Drosera. При этом для каждого растения не реже одного раза в сутки подсчитывали общее количество листьев, количество листьев с находящимися на них насекомыми и измеряли ширину наибольшего листа, а также расстояние от растения до границы распространения данной группы особей (так называемой границы популяции). По указанным выше параметрам провели также разовый обмер 25 растений из других групп особей. В ходе обработки данных наблюдений вычислили "коэффициент улавливаемости" К = n / N, где n - количество листьев с прилипшими к ним насекомыми, а N - общее количество листьев на растении. Эта величина может принимает значения от 0 до 1 и является показателем ловчей активности каждого растения.
В 2000 году из популяций Drosera anglica, D. rotundifolia и D. x obovata, расположенных на каменистом берегу озера Верхнее Пулонгское, пометили по 15 растений. Кроме того, вели контрольные наблюдения за 5 лишенными меток растениями из популяции D. rotundifolia. Из популяции D. rotundifolia, расположенной на сплавине у берега озера Левое (в 3 км к северу от деревни Н. Пулонга) также пометили 15 растений. Из всех 4 популяций было выделено по одному модельному растению. Ежедневно для каждого листа модельного растения отмечали количество насекомых, состояние пластинки, ее длину и ширину, длину черешка, а также возрастной тип листа (см. табл. 1). Наблюдения за растениями из этих популяций вели в течение 10 дней. Кроме того, пометили 10 растений вида D. rotundifolia из пятой популяции, расположенной на острове Иваньков (губа Кив), за которыми наблюдали в течение 7 дней.
Таблица 1. Возрастные типы листьев.
| тип листьев | характеристика состояния листьев |
| развивающиеся | еще не развернувшиеся листья, не способны ловить насекомых |
| активные | зрелые листья, способны ловить насекомых |
| отмершие | старые листья с нефункционирующими слизевыми железками, не способны ловить насекомых |
У всех растений один раз в сутки подсчитывали количество насекомых, находящихся на каждом активном листе, и общее количество листьев на растении. Кроме того, один раз измерили длину и ширину самого крупного листа, длину его черешка, диаметр листовой розетки и расстояние от растения до границы популяции, которой оно принадлежит.
В течение 12 дней ежедневные наблюдения проводились также за 15 растениями из популяции Pinguicula vulgaris, расположенной на каменистом берегу озера Нижнее Пулонгское. За 15 растениями из второй популяции популяции этого вида, расположенной среди мха на каменистом субстрате рядом с тропинкой на мысе Иванов Наволок, наблюдали в течение 6 дней. Один раз в сутки отмечали количество отмерших, активных и развивающихся листьев, количество насекомых на каждом активном листе, а также измеряли длину и ширину наибольшего листа, состояние его листовой пластинки, а при наличии соцветия - длину цветоноса.
Количество насекомых на листе зависело не только от интенсивности ловли и переваривания, но также и от других факторов - мы наблюдали случаи клептопаразитизма, когда муравьи уносили прилипших к листьям мертвых, но еще не переваренных насекомых.
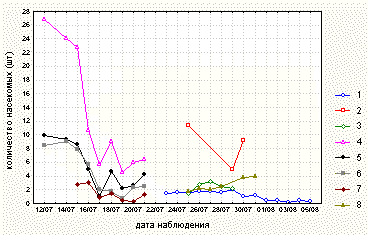
Полученные в ходе наблюдений данные обработали при помощи пакета STATISTICA for Windows (StatSoft, Inc., 1995). Для всех данных наблюдений вычислили коэффициенты корреляции. Анализировали параметрические коэффициенты корреляции, так как распределение данных было близким к нормальному. Для каждой изучаемой в 1999 и 2000 годах популяции построили графики зависимости среднего количества насекомых на растении (среднего арифметического количества насекомых на всех исследуемых растениях за одно наблюдение) и среднего количества активных листьев на растении (среднего арифметического количества активных листьев на всех исследуемых растениях за одно наблюдение) от времени наблюдения (рис. 1, 2).
В период наблюдений в 1999 году и 2000 году ветер был переменным, а средняя дневная температура воздуха и количество осадков существенно различались. В 1999 году средняя дневная температура воздуха составляла около +20°C, было пасмурно (средняя облачность около 80%), часто шел дождь. В 2000 году средняя дневная температура воздуха составляла около +25°C, небо было ясное (средняя облачность около 0%), дождя не было.
Корреляционный анализ данных, полученных в 2000 году при разовом обмере растений выявил положительную корреляцию между длиной и шириной листа у всех исследованных видов. У всех росянок обнаружили также положительную корреляцию между длиной черешка наибольшего листа и диаметром листовой розетки, тогда как для Pinguicula vulgaris эти показатели не анализировали вследствие незначительной длины черешка листа.
При анализе коэффициентов корреляции данных, полученных при ежедневных наблюдениях за Pinguicula vulgaris на берегу озера Нижнее Пулонгское видно, что количество активных листьев у Pinguicula vulgaris коррелирует с количеством развивающихся листьев. Таким образом, при увеличении общего количества листьев соотношение между количеством листьев разных типов у Pinguicula vulgaris остается прежним.
По результатам корреляционного анализа данных, полученных в ходе ежедневных наблюдений за Drosera rotundifolia и за Pinguicula vulgaris, вероятность наличия соцветия, а в случае его наличия и длина цветоноса, положительно коррелируют с длиной и шириной листьев. Отсюда можно сделать вывод, что у Drosera rotundifolia и у Pinguicula vulgaris успешность цветения повышается с увеличением размеров растения.
По данным разового обмера популяции Drosera rotundifolia на озере Верхнее Пулонгское видно, что растения с метками не отличаются по успешности ловли (то есть по общему количеству пойманных насекомых; среднему количеству насекомых на лист; количеству листьев с насекомыми и их отношению к общему количеству листьев) от растений, лишенных меток. Отсюда можно сделать вывод, что метки не оказывают существенного влияния на жизнедеятельность растений.
В ходе ежедневных наблюдений отмечали изменение количества пойманных растениями насекомых и количества разных типов листьев на растении с течением времени (рис. 1, 2).
Рисунок 1. Среднее количество насекомых на исследуемых насекомоядных растениях по данным 1999 и 2000 годов.
Условные обозначения (популяции):
1 - Pinguicula vulgaris на берегу озера Нижнее Пулонгское (по данным 2000 года)
2 - Pinguicula vulgaris на мысе Иванов Наволок (по данным 2000 года)
3 - Drosera rotundifolia на мысе Иванов Наволок (по данным 1999 года)
4 - Drosera anglica на берегу озера Верхнее Пулонгское (по данным 2000 года)
5 - Drosera x obovata на берегу озера Верхнее Пулонгское (по данным 2000 года)
6 - Drosera rotundifolia на берегу озера Верхнее Пулонгское (по данным 2000 года)
7 - Drosera rotundifolia на берегу озера Левое (по данным 2000 года)
8 - Drosera rotundifolia на острове Иваньков (по данным 2000 года)
Рисунок 2. Среднее количество активных листьев на исследуемых насекомоядных растениях по данным 1999 и 2000 годов.
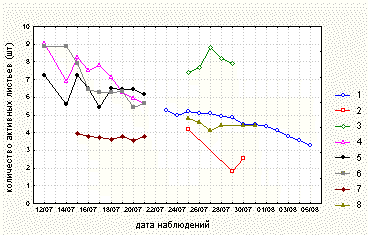
Условные обозначения: см. условные обозначения к рис. 1.
В Швеции у Drosera rotundifolia количество отмерших листьев составляло в среднем 1,3-2,5 на растение, а общее количество листьев на растении варьировало от 1 до 10 и составляло в среднем 4,5 листа на растение (Redbo-Torstensson, 1994). Эти показатели не отличаются достоверно от полученных нами для изученных нами растений.
В 1999 году успешность ловли и количество активных листьев у Drosera rotundifolia в период наблюдений увеличивались. В 2000 году у растений из всех исследуемых популяций Drosera на берегу озера Верхнее Пулонгское и у Pinguicula vulgaris на берегу озера Нижнее Пулонгское успешность ловли и количество активных листьев, наоборот, уменьшались. У растений из популяции Drosera rotundifolia на берегу озера Левое эффективность ловли насекомых также снижалась, при этом количество активных листьев в период наблюдений оставалось постоянным. У D. rotundifolia на острове Иваньков эффективность ловли в период наблюдений возрастала, а количество активных листьев уменьшалось (рис. 1, 2). Ловля насекомых у Pinguicula vulgaris на мысе Иванов Наволок происходила значительно успешнее, чем на берегу озера Нижнее Пулонгское. При этом растения Pinguicula vulgaris на мысе Иванов Наволок имели в среднем больше активных листьев, чем растения из другой популяции (табл. 2).
Таблица 2. Успешность ловли насекомых, количество активных листьев у изученных насекомоядных растений и условия их обитания по данным 1999 и 2000 годов.
| популяция | влажность субстрата | защищенность от ветра | плодородность субстрата | среднее кол-во насекомых на растение | среднее кол-во активных листьев на растении |
| Drosera rotundifolia на мысе Иванов Наволок (1999 год) | средняя | высокая | средняя | 2,4 | 8,0 |
| D. anglica на берегу озера Верхнее Пулонгское (2000 год) | низкая | низкая | высокая | 12,8 | 7,0 |
| D. x obovata на берегу озера Верхнее Пулонгское (2000 год) | низкая | низкая | высокая | 5,3 | 6,4 |
| D. rotundifolia на берегу оз. Верх. Пулонгское (2000 год) | низкая | низкая | высокая | 4,5 | 6,9 |
| D. rotundifolia на берегу озера Левое (2000 год) | высокая | средняя | низкая | 1,5 | 3,7 |
| D. rotundifolia на острове Иваньков (2000 год) | средняя | низкая | средняя | 2,7 | 4,5 |
| Pinguicula vulgaris на берегу озера Нижнее Пулонгское (2000 год) | средняя | средняя | высокая | 1,1 | 4,5 |
| P. vulgaris на мысе Иванов Наволок (2000 год) | низкая | средняя | высокая | 8,5 | 2,8 |
Как видно из этого описания, успешность ловли растениями насекомых и количество активных листьев на растениях изменялись со временем. Мы предполагаем, что успешность ловли насекомых у Pinguicula vulgaris и всех исследованных видов Drosera зависит от погодных условий (в том числе от направления ветра). Зависимость количества активных листьев от времени у D. rotundifolia в 1999 и 2000 годах различна, а в пределах одного года у всех изученных видов Drosera и Pinguicula vulgaris варьирует в зависимости от местообитания. Можно предположить, что количество активных листьев на растении у Pinguicula vulgaris и всех исследованных видов Drosera зависит как от погодных условий (в основном от количества осадков), так и от условий обитания. При этом прослеживается четкая зависимость количества активных листьев у D. rotundifolia от плодородности субстрата, тогда как изменение открытости для ветра и влажности местообитания в пределах одного года не влияют существенно на количество активных листьев и успешность ловли.
Корреляционный анализ данных, полученных в ходе разового обмера росянок, показал, что при увеличения расстояния от растения до границы популяции у D. rotundifolia возрастает длина черешка, при этом для двух других изучаемых видов Drosera влияния удаленности растения от границы популяции на размер растения не наблюдали. У D. anglica и D. x obovata при удалении растения от границы популяции уменьшается количество насекомых на листьях, тогда как у D. rotundifolia такой зависимости не обнаружили. Полученные данные позволяют говорить о наличии у всех изученных видов Drosera внутрипопуляционной конкуренции. У D. rotundifolia при горизонтальном расположении листьев удаленность от границы популяции компенсируется увеличением ловчей поверхности и не влияет на успех ловли, а приподнятые от земли листья Drosera anglica и D. obovata "фильтруют" воздушный поток и осаждают насекомых по мере их продвижения к центру популяции, поэтому у этих последних видов внутрипопуляционная конкуренция выражена особенно ярко.
Корреляционный анализ данных, полученных в ходе наблюдений за модельными растениями и разового обмера Drosera, показал взаимосвязь между количеством пойманных насекомых и показателями размера растения (то есть количеством листьев, их шириной, длиной, длиной черешка и диаметром листовой розетки). В свою очередь, количество пойманных насекомых должно зависеть от площади ловчей поверхности. Для Pinguicula vulgaris влияния количества пойманных насекомых на размерные характеристики растения не наблюдалось.
Для всех изученных видов выявлена также следующая зависимость: чем больше растение имеет листьев и чем они крупнее, тем менее равномерно пойманные насекомые распределены по листьям. Этот результат объясняется, на наш взгляд, тем, что при увеличении числа и размера листьев верхние в розетке листья начинают перекрывать нижние. В результате большинство насекомых попадает на верхние листья, а площадь ловчей поверхности нижних листьев резко сокращается.
Сравнение успешности ловли насекомых у трех исследованных видов Drosera дало следующие результаты. Наиболее успешно ловит насекомых D. anglica - она имеет наибольшие значения количества насекомых на растение, количества насекомых на лист, количества листьев с насекомыми и отношения количества листьев с насекомыми к общему количеству листьев, при этом площадь ловчей поверхности (длина и ширина листа, длина черешка, диаметр розетки) D. anglica заметно больше, чем у D. rotundifolia и D. x obovata. Последние два вида практически по всем параметрам ловли насекомых сходны друг с другом.
При анализе данных, полученных в ходе ежедневных наблюдений за модельными растениями, видно, что при сгибании листа у D. x obovata уменьшается длина листа, у D. anglica - длина и ширина листа, а также длина черешка, у D. rotundifolia - ширина листа и длина черешка. Это значит, что при поимке насекомого у D. x obovata лист сгибается в основном поперек, у D. x anglica - вдоль и поперек, а у D. x rotundifolia - в основном вдоль, причем у D. anglica и D. rotundifolia при сгибании листа изгибается также и черешок.
Таким образом, на основании полученных результатов мы можем сделать следующие
1. Интенсивность цветения у всех изученных видов насекомоядных растений зависит от их размера.
2. Успех ловли насекомых у насекомоядных растений зависит от погодных условий.
3. Количество листьев у насекомоядных растений зависит от погодных условий и от условий обитания.
4. У всех изученных видов Drosera существует внутрипопуляционная конкуренция.
5. Обнаружена связь между размером всех изученных видов Drosera и количеством пойманных растением насекомых.
6. Для Pinguicula vulgaris не выявлено влияния количества пойманных насекомых на характеристики растения.
7. Из всех изученных видов росянки Drosera anglica ловит насекомых наиболее успешно.
Работа производилась в рамках Беломорской экспедиции Московской Гимназии на Юго-Западе. В полевых наблюдениях принимали активное участие ученики и преподаватели гимназии, которым мы выражаем искреннюю благодарность.
Дарвин Ч. Насекомоядные растения // Собрание сочинений Чарльза Дарвина. Т. 4, Ч. 2. М., 1908. С. 145-154.
Баландин С.А., Баландина Т.П. Росянка круглолистная // Биологическая флора Московской области. Вып. 9, Ч. 2. М., 1993. С. 31-38.
Денисова Г.А. Росянковые // Жизнь растений. Т.5, Ч.2. М., 1981. С. 171-175.
Пельцих Л. К биологии росянки // Природа. 1917. Вып. 11-12. С. 1152-1154.
Муравник Л.Е. Ультраструктура слизистых железок Pinguicula vulgaris (Lentibulariaceae) в ходе их развития и функционирования // Бот. журн., 1988. Т. 73. N 11. С. 1523-1535.
Холодный Н.Г. Чарльз Дарвин и современные знания о насекомоядных растениях // Ч. Дарвин. Сочинения. Т. 7. М.; Л., 1948. С. 255-304.
Aldenius J., Carlsson B., Karlsson S. Effects of insect trapping on growth and nutrient content of Pinguicula vulgaris L. in relation to the nutrient content of the substrate // New Phytologist. 1983. Vol. 93. P. 53-59.
Brewer J.S. Effects of competition and litter on a carnivorous plant, Drosera cappilaris (Droseraceae) // American Journal of Botany. 1998. Vol. 85. N 11. P. 1592-1596.
Chandler G.E., Andrson J. W. Studies on the nutrion and growth of Drosera species with reference to the carnivorous habit // New Phytologist. 1976. Vol. 76. P. 129-141.
De Ridder F., Dhondt A.A. A positive correlation between naturally captured prey, growth and flowering in Drosera intermedia in two contrasting habitats // Belg. Journ. Bot. 1992. Vol. 125. N 1. P. 33-40.
Juniper B.E. Carnivorous plants. L., 1989. 353 p.
Karlsson P.S. Seasonal pattern of biomass allocation in flowering and nonflowering specimens of three Pinguicula species // Can. J. Bot. 1986. Vol. 64. P. 2872-2877.
Karlsson P.S., Carlsson B. Why does Pinguicula vulgaris L. trap insects? // New Phytologist. 1984. Vol. 97. P. 25-30.
Krafft C.C., Handel S.N. The role of carnivory in the growth and reproduction of Drosera filiformis and D. rotundifolia // Bulletin of the Torrey Bot. Club. 1991. Vol. 118. N 1. P. 12-19.
Masing V. Huulheum - Darvin Lemmikmaim // Eesti loodus. 1959. N 6.
Molau U. Reproductive ecology of the three Nordic Pinguicula species (Lentibulariaceae) // Nord. J. Bot. 1993. Vol. 13. P. 149-157.
Redbo-Torstensson P. The demographic consequences of nitrogen fertilization of a population of sundew, Drosera rotundifolia.// Acta Bot. Neerl. 1994. Vol. 43. N 2. P. 175-188.
Small J.G.C., Onraet A., Grierson D.S., Reynolds G. Studies on insect-free growth, development and nitrate-assimilating enzymes of Drosera aliciae hamet // New Phytologist. 1977. Vol. 79. P. 127-133.
Wood C.E. Evidence for the hybrid origin of Drosera anglica // Rhodora. 1955. Vol. 57. P. 105-130.
Worley A.C., Harder L.D. Consequences of preformation for dynamic resource allocation by a carnivorous herb, Pinguicula vulgaris (Lentibulariaceae) // Amer. Journ. Bot. 1999. Vol. 86. N 8. P. 1136-1145.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |