Типичный чашелистик
Типичный лепесток
Типичная тычинка

В морфологии растений наиболее распространена концепция «железной триады» органов, в соответствии с которой многообразие органов цветковых растений принято сводить к трем основным типам: корень, стебель (осевые структуры) и лист (аппендикулярный орган). Согласно фолиарной концепции цветка Гете (Arber, 1946) органы цветка также представляют собой видоизмененные филломы. Эта концепция послужила основой для разработки АВС-модели генетической регуляции типа органа в цветках (Coen, Meyerowitz, 1991), постулирующей, что «начальным» типом органа является вегетативный лист, тогда как аппендикулярные органы цветка образуются после активизации генетических функций А, В и/или С, что приводит к изменению программы развития филлома и формированию соответствующего органа. Заметим, что все многообразие органов в АВС-модели исчерпывается вегетативными листьями, чашелистиками, лепестками, тычинками и карпеллами. Таким образом, в основе преобладающих в современной науке подходов лежит априорное допущение о существовании четко разграниченных типов структурных элементов и возможности однозначно отнести каждую отдельную структуру к одному из них (Timonin, 2002). Эти четко очерченные типы структурных элементов С.В. Мейен (1978) предолжил называть меронами.
Однако у высших растений встречаются органы с промежуточными по отношению к традиционно выделяемым меронам морфологическими признаками. Подобные переходные структуры встречаются как в вегетативной так и в генеративной сфере растений. Так филлокладии Asparagaceae, модули ряда видов Utricularia (Lentibulariaceae), различающиеся только длительностью роста, степенью разветвленности и наличием ловчих пузырьков, филломорфы некоторых видов Streptocarpus обладают как морфологическими признаками оси, так и признаками листа. Последние представляют собой ось, переходящую терминально в листовую пластинку и несущую на верхней стороне такие же филломорфы и соцветия (Timonin, 2002). Классическим примером переходных структур в генеративной сфере, приведенным еще в «Метаморфозе растений» Гете (Arber, 1946), являются органы в цветках некоторых видов кувшинки (Nymphaea, Nymphaeaceae), промежуточные по морфологии между лепестками и тычинками. К видам, у которых наблюдается постепенный переход от органов околоцветника к андроцею, можно отнести широко распространенную в Европе N. alba L. s.l. (Heslop-Harrison, 1955; Дубына, 1982).
Цветки N. alba одиночные, крупные, сидящие на длинном цветоносе, обоеполые актиноморфные с двойным околоцветником. Чашелистики снизу зеленые, сверху зеленовато-белые числом от 3 до 7, как правило, 4. Лепестки белые многочисленные. Тычинки также многочисленные, располагаются по спирали, примитивного типа: тычиночные нити лентовидные с хорошо выраженным надсвязником. Пыльники очень длинные, погруженные в стерильную ткань тычинок. Гинецей синкарпный из 5-35 кондупликатных плодолистиков, с сидячим рыльцем (Heslop-Harrison, 1955; Schneider et al., 2003). Помимо широко известного существования переходных по морфологии органов между лепестками и тычинками, интересным, не отмеченным ранее в литературе фактом, является наличие переходных между чашелистиками и лепестками структур. Степень плавности перехода между лепестками и тычинками считается важным диагностическим признаком в роде Nymphaea (см. например, Uotilla, 2001), однако до сих пор не было предпринято попыток количественно охарактеризовать этот признак. Описание структур цветка кувшинки с позиций классической морфологии не отражает всего их разнообразия (Sattler, 1996; Тимонин, 2001). Следовательно, необходимо выработать иной подход, позволяющий адекватно исследовать специфику переходных структур.
Для систематиков существование растений, сочетающих в себе признаки нескольких таксонов -- их обычно считают межвидовыми гибридами (Wilson, 1992), является общепризнанным фактом. Возможно, именно поэтому основные методы, позволяющие эффективно описать морфологию переходных структур, могут быть заимствованы морфологами именно из систематики.
Одним из таких подходов является динамическая морфология, предложенная R. Sattler (1966). Основной идеей динамической морфологии является замена жесткой системы типов органов на количественную (например, балльную) оценку степени проявления у них тех или иных признаков. Такой подход в точности соответствует методу вычисления гибридных индексов для отдельных растений, предложенному для изучения интрогрессии E. Anderson в 1949 году (Wilson, 1992).
Другим подходом, используемым в систематике гораздо шире метода гибридных индексов, является применение разнообразных методов вычисления сходства между отдельными растениями после выяснения их морфологических характеристик (Wilson, 1992; Sattler, Rutishauser, 1997), основанные на вычислении разных типов дистанций и других методов многомерной статистики (Lacroix et al., 2003).
Методы динамической морфологии неоднократно применялись для исследования переходов между разными типами вегетативных органов (корень, стебель, лист – см. Sattler, 1996), тогда как переходные структуры в цветке, насколько нам известно, впервые рассматриваются с позиций динамической морфологии только в настоящей статье. Исследования динамической морфологии цветка представляют большой интерес в свете появившейся в последнее время ABC-модели развития цветка (Coen, Meyerowitz, 1991).
В июне-сентябре 2004 года мы исследовали цветки N. alba L. s.l. incl. N. candida Presl. (Uotilla, 2001) из четырех регионов Европейской России: Мосальский р-н Калужской обл. (три цветка), Удомельский (11 цветков) и Вышневолоцкий (два цветка) р-ны Тверской обл., Лоухский р-н респ. Карелия (озера Спелое и Эврика; 7 цветков) и Кизильский р-н Челябинской обл. (два цветка).
У каждого из 25 исследованных цветков мы фиксировали основные морфологические признаки всех его органов, продвигаясь последовательно от периферии цветка к его центру: (1) качественые (табл. 1) и (2) количественные (длина и ширина органа, длина его пыльниковой части и расстояние от основания органа до его наибольшей ширины). Мы отмечали цвет дистальной абаксиальной части органа по визуальной девятибалльной шкале. Пыльниковой частью органа мы называем дистальную часть тычинки, на которой располагаются пыльники, а в случае отсутствия пыльников -- геометрически обособленную дистальную часть переходного органа. Всего было исследовано 2479 органов цветка.
|
признак |
“шкала A” |
“шкала B” |
||
|
значение признака |
балл |
значение признака |
балл |
|
|
расположение пыльников |
пыльники отсутствуют |
1 |
пыльники отсутствуют |
1 |
|
под острым углом друг к другу |
7,5 |
под острым углом друг к другу |
5 |
|
|
практически параллельно |
10 |
практически параллельно |
10 |
|
|
цвет органа |
коричневый |
1 |
--- |
--- |
|
зелено-коричневый |
2 |
--- |
--- |
|
|
зеленый |
3 |
--- |
--- |
|
|
бело-зеленый |
4 |
бело-зеленый |
1 |
|
|
белый |
5 |
белый |
2 |
|
|
светло-желтый |
7 |
светло-желтый |
5 |
|
|
грязно-желтый |
8 |
грязно-желтый |
6 |
|
|
желтый |
9 |
желтый |
8 |
|
|
желто-оранжевый |
>
10 |
желто-оранжевый |
10 |
|
|
отношение длины органа к его ширине |
[1,4; 3] |
1 |
[1,4; 2,4] |
1 |
|
(3; 4] |
6 |
(2,4; 3,2] |
3 |
|
|
(4; 5] |
7,5 |
(3,2; 5,0] |
5 |
|
|
(5; 6] |
8,5 |
(5,0; 10,0] |
7 |
|
|
(6; 22] |
10 |
(10,0; 22,0] |
10 |
|
|
относительная длина пыльниковой части органа |
0 |
1 |
0 |
1 |
|
(0; 0,1] |
6 |
(0; 0,2] |
3 |
|
|
(0,1; 0,25] |
7,5 |
(0,2; 0,3] |
5 |
|
|
(0,25; 0,4] |
8,5 |
(0,3; 0,4] |
7 |
|
|
(0,4; 0,7] |
10 |
(0,4; 0,7] |
10 |
|
|
положение наибольшей ширины |
--- |
--- |
(0,6; 0,9] |
1 |
|
(0,4; 0,6] |
3 |
|||
|
(0,3; 0,4] |
5 |
|||
|
(0,1; 0,3] |
7 |
|||
|
[0; 0,1] |
10 |
|||
Известно, что условия произрастания кувшинки и возраст растений существенно влияют на размерные характеристики кувшинки, в том числе на размер цветков и их отдельных органов (Heslop-Harrison, 1955; Дубына, 1982). Для того чтобы избежать влияния размерных характеристик на наше восприятие морфологии органа, мы ввели ряд морфологических индексов (табл. 1). Кроме того, мы вычислили ряд интегральных характеристик органов на основании их основных качественных и относительных количественных морфологических признаков.
Для каждого цветка мы вычислили медианные значения всех морфологических характеристик для всех тычинок данного цветка, которые были использованы в качестве морфологических характеристик «типичной тычинки». Для каждого органа мы вычислили степень его отличия от «типичной тычинки» как эвклидово расстояние между данным органом и «типичной тычинкой», вычисленное по совокупности морфологических признаков.
Каждый орган мы охарактеризовали баллом от 1 до 10. Мы разработали две шкалы: (1) «шкала A», где типичному чашелистику соответствовал балл 1, а типичной тычинке – балл 10 и (2) «шкала B», где типичному лепестку соответствовал балл 1, типичной тычинке балл 10, а чашелистики и переходы от чашелистиков к лепесткам были исключены из анализа. Значения морфологических характеристик были переведены в баллы согласно табл. 1. Мы не переводили в баллы «шкалы A» положение наибольшей ширины, так как ее типичные значения для чашелистиков находятся между типичных значений для лепестков и для тычинок (табл. 2).
|
тип органа |
расположение пыльников |
цвет органа |
dl/sh |
pyl/dl |
полож. наиб. ширины |
|
чшл |
пыльники отсутствуют |
коричневый, зелено-коричневый или зеленый |
1,9-2,4 |
0-0 |
0,4-0,5 |
|
чшл/лп |
пыльники отсутствуют |
бело-зеленый |
1,9-2,2 |
0-0 |
0,5-0,6 |
|
лп |
пыльники отсутствуют |
белый |
1,9-2,4 |
0-0 |
0,5-0,6 |
|
лп/тч |
под острым углом друг к другу |
светло-желтый |
3,2-5,0 |
0,2-0,4 |
0,3-0,4 |
|
тч |
практически параллельно |
грязно-желтый, светло-желтый, желтый или желто-оранжевый |
6,0-10,0 |
0,3-0,4 |
0-0,4 |
Для каждого органа по каждой шкале вычислялся итоговый балл как среднее арифметическое баллов по соответствующей шкале для всех морфологических характеристик этого органа (табл. 3).
|
значение балла |
число органов с соответствующим значением балла |
|
|
по “шкале A” |
по “шкале B” |
|
|
[1;2) |
611 |
489 |
|
[2;3) |
17 |
28 |
|
[3;4) |
22 |
16 |
|
[4;5) |
0 |
142 |
|
[5;6) |
39 |
268 |
|
[6;7) |
10 |
382 |
|
[7;8) |
220 |
595 |
|
[8;9) |
707 |
375 |
|
[9;10] |
853 |
52 |
Влияния размерных характеристик на восприятие формы органа также позволяет избежать геометрическая морфометрия, которая анализирует форму объекта как таковую, без учета его размерных характеристик (Adams et al., 2002). Наиболее распространенной группой методов геометрической морфометрии являются методы, которые используют постановку на исследуемом контуре точек, отмечающих его ключевые места (т.н. «меток»), такие как Thin Plate Splines analysis (TPS). Эти метки должны быть гомологичны хотя бы геометрически, так как методы, основанные на метках, оперируют только с их координатами, и различия между объектами должны быть сравнимыми. TPS вычисляет степень деформации, необходимую для совмещения меток на одном образце с соответствующими метками другого образца -- прокрустова трансформация (Adams et al., 2002). Мерой различия между формами по координатам меток служит прокрустово расстояние, которое является метрикой, с которой при обработке данных можно обращаться так же, как с эвклидовым расстоянием (Pavlinov, 2001). К результатам TPS могут быть применены многомерные методы анализа данных, так как проекция каждой формы на плоскость двух первых главных компонент, вычисленных из матрицы прокрустовых трансформаций, может играть роль обычной матрицы данных (Bookstein, 1991).
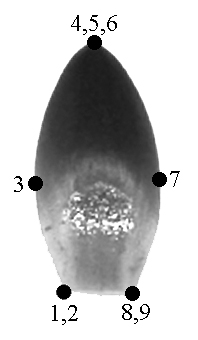
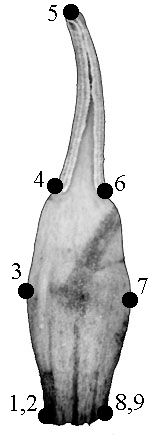
Для того, чтобы протестировать возможность применения методов геометрической морфометрии к нашему объекту, мы обвели на бумаге каждый из 73 органов одного цветка из Кизильского р-на Челябинской обл. (далее модельный цветок). Для характеристики формы органа мы использовали 9 меток, расположенных на его контуре. В нашем случае для фиксации меток использовались не только биологические, но и геометрические соображения: метки расставлялись в точках наибольшей кривизны контура (рис. 1). Координаты точек снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig (Rohlf, 2001b). Координаты эталонной конфигурации, а также значения главных, относительных и частных трансформаций, характеризующих меру отличия образца от эталона (Bookstein, 1991), вычисляли при помощи программы tpsRelw (Rohlf, 2001a). Исходные координаты экземпляров нормировали относительно эталонной конфигурации при помощи прокрустова наложения, коэффициент шкалирования альфа принимали равным 0. Прокрустовы расстояния вычисляли при помощи программы tpsSplin (Rohlf, 1997).
|
Типичный чашелистик |
Типичный лепесток |
Типичная тычинка |
|
|
|
При построении кривых изменения различных характеристик органов цветка при продвижения от периферии цветка к его центру мы использовали метод кубического сглаживания, который наиболее адекватен при сложной конфигурации кривых. Все вспомогательные вычисления, все виды графического и многомерного анализа матриц значений частных деформаций и шкалированных результатов морфологических промеров (многомерное шкалирование, анализ главных компонент, дисперсионный анализ, дискриминантный анализ), а также сопоставление матриц прокрустовых расстояний и эвклидовых расстояний (мантель-тест, использовано 1000 пермутаций) и корреляционный анализ данных были проведены при помощи статистической среды R 1.9.1 (R Development Core Team, 2004).
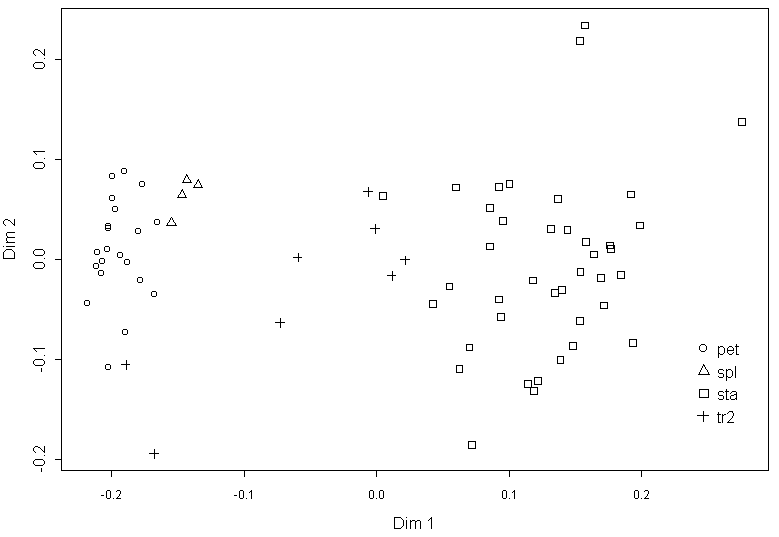
В процессе измерений мы относили каждый орган к одному из пяти типов (чашелистик, переход от чашелистика к лепестку, лепесток, переход от лепестка к тычинке, тычинка) на основании классического подхода (Heslop-Harrison, 1955; Schneider et al., 2003). Типичные морфологические характеристики указанных органов приведены в табл. 2. Отнесение органов цветка кувшинки к одному из пяти выделенных нами классов (три основных и два переходных) на основании их внешнего вида с очень высокой степенью достоверности соответствует результатам классификации этих органов по морфологическим признакам. Об этом свидетельствуют результаты многомерного дисперсионного анализа (MANOVA: p<0,001) и дискриминантного анализа (Wilks lambda 0,02, p<0,001, misclassification error 22%), результаты анализа главных компонент представлены на рис. 2, результаты многомерного шкалирования и анализа главных компонент для всех остальных цветков имеют похожий вид (не приведены).
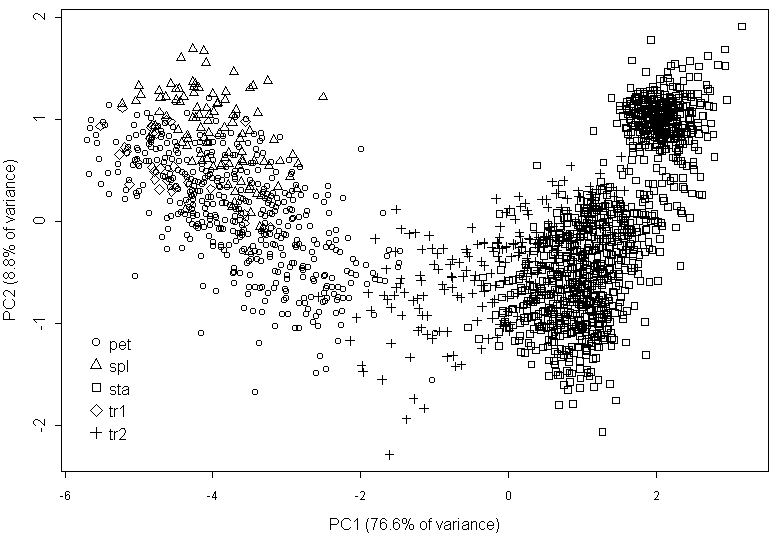
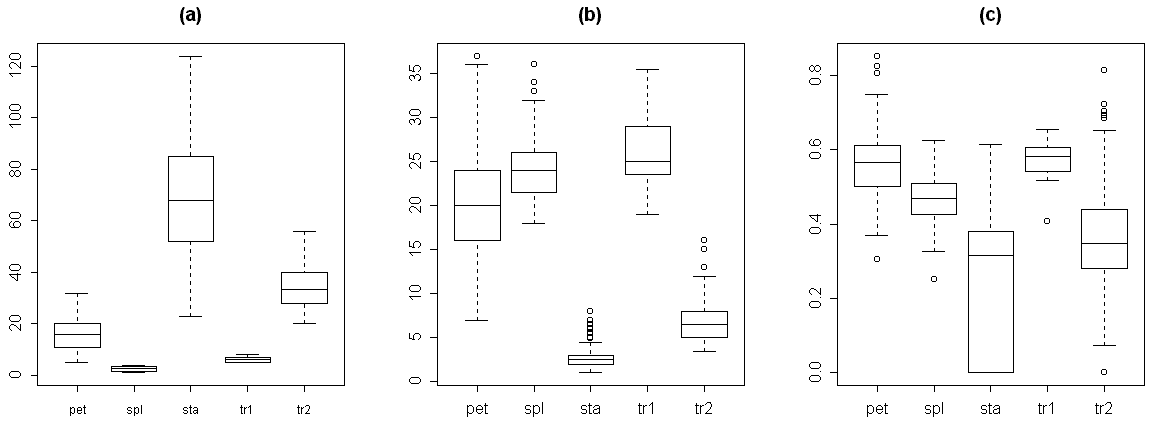
Условные обозначения: тип органа по визуальной оценке: spl -- чашелистики; tr1 -- переходные между чашелистиками и лепестками органы; pet -- лепестки; tr2 -- переходные между лепестками и тычинками органы; sta -- тычинки
Переходные между чашелистиками и лепестками структуры по морфологии чрезвычайно близки к чашелистикам (рис. 2). Основным морфологическим является наличие хлорофилльной полоски на дистальной части абаксиальной стороны органа при его общей белой окраске (табл. 2). Наличие в цветке нескольких (от одного до четырех) переходных между чашелистиками и лепестками структур характерно для наших образцов из Тверской области.
Переходные между лепестками и тычинками структуры по совокупности морфологических признаков четко отличаются и от лепестков, и от тычинок (рис. 2, табл. 2). Все вышесказанное позволяет нам использовать предварительную типификацию органов на основании их внешнего вида в дальнейшем анализе данных. При этом надо учесть, что 36% переходных между лепестком и тычинкой структур (судя по морфологическим признакам) было отнесено к тычинкам по их внешнему виду, а 6% тычинок – к переходным между лепестком и тычинкой структурам.
Такие расхождения могут свидетельствовать о существовании значимых для человеческого глаза признаков формы органа, которые не могут быть охарактеризованы морфологическими промерами. Поэтому мы решили сравнить результаты классификации органов цветка по морфологическим данным и по форме органа в целом (данные геометрической морфометрии). Матрица прокрустовых расстояний (данные геометрической морфометрии) и матрица эвклидовых расстояний (данные классической морфометрии) между органами модельного цветка высоко скоррелированы (мантель-тест: r=0,77, p<0,01).
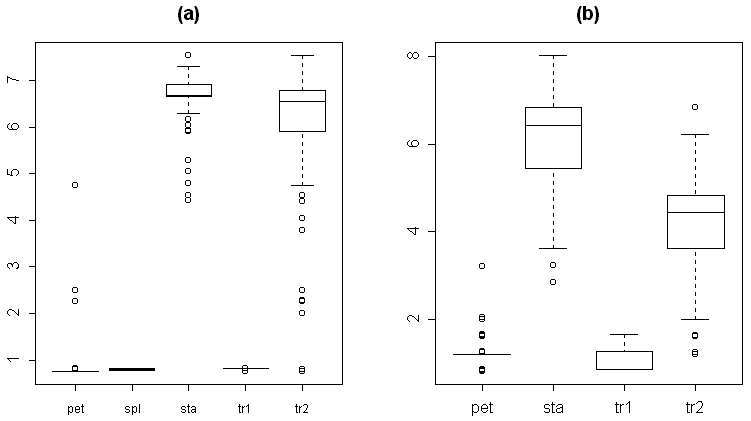
На графиках изменения значений отдельных морфологических признаков органов цветка кувшинки при продвижении от периферии цветка к его центру хорошо видно, что первый перегиб кривых приходится на разные органы, причем не всегда заранее ясно, на какие (рис. 3a, 3b). Мы постарались показать различные типы изменения морфологических признаков при продвижении от периферии цветка к его центру. Например, при смене лепестков переходными между лепестками и тычинками структурами перегиб кривой изменения ширины для кувшинок из Тверской обл. приходится на промежуток между 30-ым и 40-ым органом, что соответствует переходным между лепестком и тычинкой органам, а для кувшинок из респ. Карелия – примерно на 30-ый орган, что соответствует тычинке, (рис. 3a). Перегиб кривой изменения относительной величины пыльников приходится соответственно примерно на 22-ой (лепесток) и на 18-ый (лепесток) орган (не приведено). Перегиб кривой изменения положения наибольшей ширины органа у кувшинок из Тверской обл. слабо выражен и приходится на промежуток между 30-ым и 45-ым органом (переходные между лепестками и тычинками органы), а для кувшинок из респ. Карелия перегиб не выражен, и у всех типов органов цветка положение наибольшей ширины составляет около 0,5 (рис. 3b). Таким образом, можно разделить морфологические признаки на те, которые позволяют относительно успешно разграничивать разные типы органов (например, относительная длина пыльников и в меньшей степени ширина органа), и на те, которые не позволяют провести это разграничение сколь-нибудь успешно (например, положение наибольшей ширины органа). Из вышесказанного следует, что изменение различных морфологических характеристик органов цветка, обуславливающее в конечном итоге смену чашелистиков лепестками, а лепестков тычинками, в цветке кувшинки происходит асинхронно.
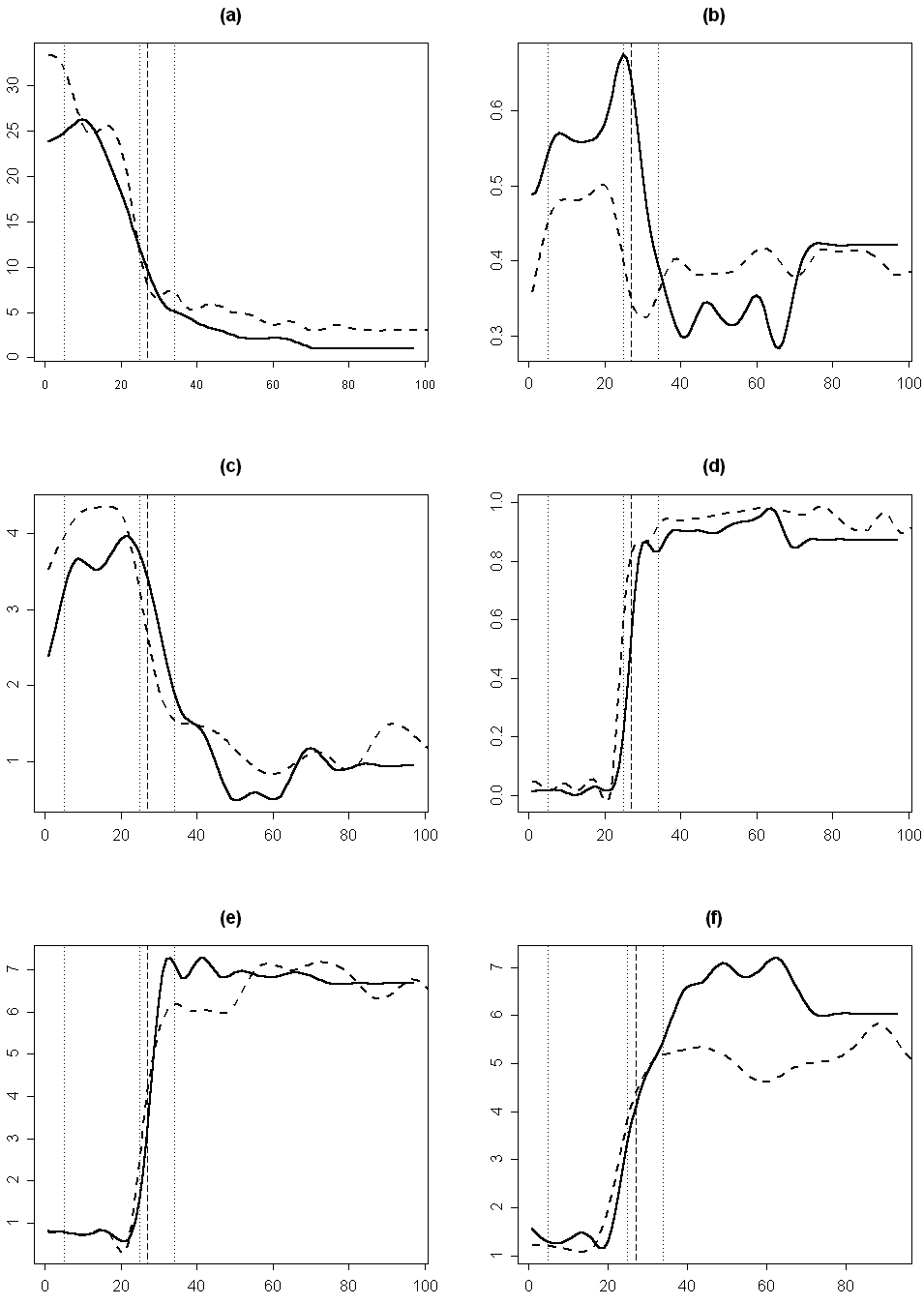
(a) наибольшая ширина органа, (b) положение наибольшей ширины органа, (c) многомерное эвклидово расстояние до "типичной тычинки", (d) вероятность принадлежности к кластеру "типичной тычинки", (e) значение индекса по "шкале А", (f) значение индекса по "шкале B". Вертикальные линии обозначают границы между разными типами органов: две первые точечные линии (слева направо) -- между чашелистиками и лепестками и между лепестками и переходными между лепестками и тычинками органами, соответственно, третья точечная линия и пунктирная линия обозначают границу между переходными между лепестками и тычинками органами и тычинками для цветка №35 (третья точечная линия) и цветка №26 (пуктирная линия).
Эвклидово расстояние (как мера несходства органа с «медианной тычинкой») несколько увеличивается при переходе от чашелистиков к лепесткам и существенно снижается при переходе от лепестков к тычинкам, не достигая при этом нуля (рис. 3c). Скорость уменьшения эвклидова расстояния при переходе от лепестков к тычинкам (выраженная через модуль производной эвклидова расстояния) обратно зависит от доли переходных между лепестками и тычинками структур в цветке (r= -0,6; p<0,01). Надо отметить, что перегиб кривой изменения эвклидова расстояния не всегда точно соответствует смене одного типа органов (выделяемого по совокупности морфологических признаков) другим. Кроме того, эта кривая, как правило, имеет довольно сложный вид, что затрудняет выделение интересующих нас перегибов (рис. 3c).
Кривые изменения баллов по «шкале A» (табл. 3) при продвижении от периферии к центру цветка имеют относительно простой вид с четкими перегибами в местах смены одного типа органов другим (рис. 3e). При продвижении от периферии цветка к его центру значения баллов по «шкале A» монотонно увеличиваются, причем наиболее резкое скачкообразное увеличение приходится на участок переходных между лепестками и тычинками структур. Для некоторых цветков этот участок графика имеет вид почти вертикальной прямой. Изучение изменения баллов по «шкале B» (табл. 3) позволяет более подробно проанализировать процесс перехода лепестков в тычинки (рис. 3f). Кривые изменения баллов по этой шкале имеют вид, чрезвычайно сходный с видом кривых изменения баллов по «шкале A». По внешнему виду кривых изменения баллов по «шкале B» четко выделяются кувшинки из респ. Карелия, у которых практически отсутствуют переходные между лепестками и тычинками структуры (0-4% от общего числа органов в цветке). Тычинки этих цветков по морфологии более сходны с лепестками по сравнению с цветками из других регионов Европейской России, для них характерны некоторые черты переходных между лепестками и тычинками органов (табл. 2). У цветков из респ. Карелия наибольшая ширина находится почти в середине тычинки (рис. 3…), длина тычинки превышает ее ширину в 4-6 раз, а пыльники могут располагаться под острым углом друг к другу, тогда как у цветков из других регионов Европейской России наибольшая ширина находится ближе к основанию тычинки, длина тычинки превышает ее ширину в 7-11 раз, а пыльники всегда параллельны друг другу.
Условные обозначения: см. усл. обозн. к рис. 2
Сравнение визуальной классификации органов цветка кувшинки на основании их внешнего вида и их классификации методом анализа главных компонент на основании морфологических признаков выявило различия между этими классификациями для тычинок и переходных между лепестками и тычинками структур. Сходство результатов, полученных методами классической и геометрической морфометрии, свидетельствует о том, что относительные морфологические показатели так же, как и методы геометрической морфометрии, успешно характеризуют форму органов как таковую. Анализ кривых изменения морфологических характеристик органов цветка кувшинки при продвижении от периферии цветка к его центру показал, что отдельные морфологические признаки не могут быть использованы для уверенного отнесения органов к тому или иному типу. Таким образом, данных классической морфологии недостаточно для описания переходных между лепестками и тычинками органов. Это значит, что для исследования подобных переходных структур мы должны использовать некий интегральный показатель, объединяющий в себе все существенные морфологические характеристики этих органов.
В качестве такого показателя могут быть использованы баллы динамической морфологии, которые оптимально характеризуют переходные структуры генеративной сферы цветковых растений, что подтверждает ранее полученные обширные результаты для вегетативных органов высших растений (Sattler, 1966; Sattler, 1996; Sattler, Rutishauser, 1997; Lacroix, 2003). Анализ баллов динамической морфологии позволил нам выявить различия в морфологии цветка кувшинок из различных регионов России, что может иметь и таксономическое значение. Таким образом, динамическую морфологию можно считать чрезвычайно эффективным методом для исследования переходных структур и в генеративной сфере цветковых растений.
Другим возможным интегральным показателем, позволяющим довольно успешно охарактеризовать переходные структуры, является эвклидово расстояние между органом и «медианной тычинкой», вычисленное на основании совокупности морфологических признаков. Надо отметить, что вычисления эвклидова расстояния требуют оперирования с n-мерным пространством, где n – это число использованных для характеристики органа морфологических признаков. Очевидно, что при n>3 использование компьютерной обработки данных становится обязательным условием применения этого подхода. Это ограничение, переставшее быть существенным недостатком в эпоху всеобщей компьтеризации, одновременно является и достоинством предлагаемого метода. Эвклидово расстояние может быть вычислено с применением несложных программ почти мгновенно вне зависимости от объема выборки и числа используемых признаков. Кластерный анализ, основанный на вычислении эвклидовых расстояний, был успешно применен C. Lacroix с соавторами (2003) для исследования структур вегетативной сферы (сопоставления морфологии побегов, сложных листьев и их листочков).
Методы вычисления эвклидовых расстояний и баллов динамической морфологии могут быть с успехом применены для исследования самых разных переходных структур генеративной сферы как в морфологии, так и в генетике. Так, предложенные методы актуальны при изучении плавных морфологических переходов, например, разных типов околоцветника у некоторых представителей сем. Cactaceae, при котором нижние органы имеют черты чашелистиков или даже вегетативных листьев, а верхние обладают признаками лепестков (!!! Волгин). Если в цветках можно выделить более 4 классических типов органов (как у Parnassia), разработанные методы позволят более определенно судить о природе «проблемных» органов и решать возникающие терминологические трудности. Наиболее широкое применение методы динамической морфологии и вычисления эвклидовых расстояний должны найти в генетике для точного описании органов мутантных растений. Например, на модельном объекте Arabidopsis thaliana (L.)Heyn. у мутантов apetala2 отмечены листовидные органы, несущие ткань рыльца и семязачатки, т.е. промежуточные между листьями и карпеллами (Bowman et al., 1989). У растений томатов с нарушенной экспрессией LeT6-гена (гомеобокс-содержащий ген томтов класса I) у сложного листа появляются признаки осевого органа (Janssen et al., 1998).
Согласно активно распространившейся в последнее время «ABC-модели» генного контроля развития цветка, дифференциация лепестков и тычинок обуславливается экспрессией генов группы B. Лепестки закладываются в тех участках, где наряду с генами группы B экспрессируются гены группы A, а тычинки – там, где наряду с генами группы B экспрессируются гены группы C (Coen, Meyerowitz, 1991; обзор современного состояния проблемы см. Soltis et al., 2002). Таким образом, смена лепестков тычинками с позиций генетики обуславливается сменой экспресии одних генов экспрессией других генов. По ABC-модели экспрессия генов каждого класса поисходит в строго очерченных доменах флорального примордия, что выражается в четкой границе между органами чашечки, венчика, андроцея и гинецея. В этом случае наиболее уместно рассматривать цветок в рамках типологического подхода.
У Nymphaea ортологи АВС-генов до сих пор слабо охарактеризованы (Soltis et al., 2002). В цветке кувшинки морфология органов изменяется постепенно, т.е. можно предположить, что границы между зонами экспрессии генов недостаточно выражены и эти зоны довольно плавно переходят друг в друга. Так, граница между андроцеем и околоцветником определяется границей экпрессии генов А и С класса (Coen, Meyerowitz, 1991). По всей видимости у кувшинок существует зона, где активны гены обоих классов, поэтому для описания органов динамический подход более адекватен. Примечательно, что границы, проведенные на основании выраженного изменения того или иного признака, часто не совпадают и приходятся на разные органы. Если предположить, что все изученные нами морфологические признаки у кушинок находятся под контролем АВС-системы, то должны существовать пороговые значения экспрессии каждого гена, влияющие на проявления признака. Причем для проявления разных признаков пороговые значения экспрессии одного и того же гена различаются. Этим можно объяснить асинхронное изменение различных морфологических характеристик органов цветка при продвижении от лепестков к тычинкам. Вполне возможно, что смена экспрессируемых групп генов в цветках других таксонов покрытосеменных растений также происходит не столь резко, как это предполагается в классической ABC-модели (Coen, Meyerowitz, 1991; Soltis et al., 2002), или контролируется дополнительными генами, не включенными в АВС-модель.
В литературе до сих пор не прекращаются споры о том, какой подход является более обоснованным: типологическая или динамическая морфология (Sattler, 1966; Sattler, Rutishauser, 1997; Тимонин, 2001 и др.). Основным возражением против динамической морфологии считается необходимость выбора эталонов проявления соответствующего признака, при сравнении с которыми и выставляется балл. Такие эталоны присущи только мерону, выявляемому в рамках типологической морфологии (Timonin, 2002). Однако такими эталонами проявления признака можно считать морфологические признаки экстремальных морфотипов, между которыми существует переход. Если мероны представить в виде таких экстремальных морфотипов, чем они в сущности и являются, то различия между типологической и динамической морфологией исчезнут (Sattler, 1996). Даже при отрицании существования переходных форм между меронами типологическая и динамическая морфология остаются взаимодополняющими, а не исключающими друг друга подходами (Sattler, 1996).
Большая часть сбора данных проводилась на Молдинской биостанции Московской Гимназии на Юго-Западе (№1543) и в ходе Беломорских экспедиций Гимназии (руководитель С.М. Глаголев). Мы искренне благодарны ученикам Гимназии Александровой О., Блохиной Н., Величанской Н., Глаголеву А., Кудиной О., Кулыгиной Н., Мордвинкину Д. и Червячковой Е. за активное участие в сборе данных и Альтшулеру Е. за своевременную помощь в программировании на языке R.
Дубына Д.В. Кувшинковые Украины. Киев. Наукова думка. 1982.
Мейен С.В. Основные аспекты типологии организмов // Ж. общ. биол. 1978. Т. 39. N 4. С. 495-508.
Adams D.C., Rohlf F.J., Slice D.E. Geometric Morphometrics: Ten Years of Progress Following the ‘Revolution’ [Electronic resource]. 2002. Mode of access:
http://life.bio.sunysb.edu/morph/review/review.html.
Arber A. Goethe’s botany. The meta-morphosis of plants (1790) and Tobler’s Ode to Nature (1782) with an intriduction and translation by Agnes Arber // Chronica Botanica. Vol. 10. P. 66-124.
Bookstein F.L. Morphometric tools for landmarks data. New York: Cambridge Univ. Press. 1991.
Bowman J.L., Smyth D.R., and Meyerowitz E.M. Genes directing flower development in Arabidopsis // Plant Cell. 1989. Vol. 1. P. 37–52.
Coen E., Meyerowitz E. The war of the whorls: genetic interactions flower development // Nature. 1991. Vol. 353. P. 31–37.
Heslop-Harrisson J. Nymphaea // J. Ecol. 1955. Vol. 43. P. 719-734.
Janssen B.-J., Lund L., and Sinha N. Overexpression of a homeobox gene, LeT6, reveales indeterminate features in the tomato compound leaf // Pl. Phys. 1998. Vol. 117. P. 771-786.
Lacroix C., Jeune B., Purcell-MacDonald S. Shoot and compond leaf comparisons in eudicots: dynamic morphology as an alternative approach // Bot. J. Linn. Soc. 2003. Vol. 143. P. 219-230.
Pavlinov I. Ja. Geometric morphometrics, a new analytical approach to comparison of digitized images // Information technology in biodiversity research. 2001. P. 41-64.
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. 2004.
Rohlf F.J. tpsSplin. N.Y.: State Univ. at Stony Brook. 1997. [Electronic resource]. Mode of access:
http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsRelw: relative warps. N.Y.: State Univ. at Stony Brook. Version 1.23. 2001a. [Electronic resource]. Mode of access:
http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsDig. N.Y.: State Univ. at Stony Brook. Version 1.23. 2001b. [Electronic resource]. Mode of access:
http://life.bio.sunysb.edu/morph/.
Sattler R. Towards a more adequate approach to a comparative morphology // Phytomorphology. 1966. Vol. 16. P. 417-429.
Satter R. Classical morphology and continuum morphology: opposition and continuum // Ann. Bot. 1996. Vol. 78. P. 577-581.
Sattler R., Rutishauser R. The fundamental relevance of morphology and morphogenesis to plant research // Ann. Bot. 1997. Vol. 80. P. 571-582.
Soltis D.E., Soltis P.S., Albert V.A., Oppenheimer D.G., Hong M., dePamphilis C.W., Frondlich M.W., Theissen G. (Floral Genome Project Research Group). Missing links: the genetic architecture of flower and floral diversification // Trends in plant science. 2002. Vol. 7. N 1. P.22-31.
Schneider E.L., Tucker S.C., Williamson P.S. Floral development in the Nymphaeales // Int. J. Plant Sci. 2003. Vol. 164(5 Suppl.). P. 279–292.
Timonin A.C. Sattler’s dynamic morphology: an acme or a reverie? // Wulfenia, 2002. Vol. 9. P. 9–18.
Uotilla P. Nymphaea L. // Flora Nordica. 2000. Vol. 2. P. 216-221.
Wilson P. On inferring hybridity from morphological intermediacy // Taxon. 1992. Vol. 41. N 1. P. 11-23.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |