Род Nymphaea широко распространен в зоне тропического и умеренного климата обоих полушарий. Кувшинки обладают не только определенной эстетической ценностью, украшая водоемы и оранжереи ботанических садов, но и издавна применяются в народной медицине. Разновидности и формы видов кувшинки имеют разные по числу, размерам и внешнему виду цветки и листья, что позволяет на основании изучения их экологических условий произрастания создавать аналогичные формы в искусственных бассейнах и рекомендовать лучшие из них для зеленого строительства. Важным вопросом является также отбор форм кувшинковых со значительным содержанием биологически активных веществ и высокой продуктивностью листьев и корневищ, а также устойчивых к загрязнению водоемов. В связи с этим изучение морфологического разнообразия представителей рода кувшинка представляет собой важную задачу.
До настоящего времени не существует единого мнения о таксономической ситуации в роде кувшинка в европейской части России. Целью настоящей работы было всестороннее изучение морфологической изменчивости кувшинки в указанном регионе.
Род Nymphaea содержит около 40-50 видов, распространенных как в тропической зоне, так и в зоне умеренного климата обоих полушарий. Во многих видах выделяют многочисленные подвиды, хромосомные расы и многочисленные формы гибридного и искусственного происхождения. Эти виды имеют широкую экологическую амплитуду, обитая на минеральных или органических отложениях в водоемах с реакцией среды от кислой до щелочной. В России произрастает три вида кувшинки: N. alba L., N. candida J. Presl. и N. tetragona Georgi. N. alba распространена в Европейской части России, N. candida – в Европейской части России и в Сибири, N. tetragona – в Сибири, на Дальнем Востоке и на Кольском полуострове (Комаров, 1937; Muntendam et al., 1996). Надо отметить, что П. Уотилла, допуская возможность видовой самостоятельности N. alba и N. candida, придает им ранг подвида (N. alba subsp. alba и N. alba subsp. candida соответственно) “из практических соображений” (Uotilla, 2001).
До настоящего времени не существует более или менее общего мнения по поводу трактовки видов рода Nymphaea, в частности N. alba, N. candida и N. tetragona (Heslop-Harrison, 1955; Маевский, 1964; Колесниченко, 1977).
Полиплоидия, хромосомные перестройки и генные мутации сыграли важную роль в эволюции кувшинок (Wiersema, 1988). Например, для N. alba известны следующие хромосомные числа: 2n=64, 96, 48, 84, 105, 112 (Heslop-Harisson, 1955), 52, 56, 160 (Крупкина, 2001), 108 (Дубына, 1982). Основным хромосомным числом для рода Nymphaea считают 7, 8 или 14. По-видимому, точное определение хромосомных чисел затрудняется большим числом и мелкими размерами хромосом (Heslop-Harrison, 1955). На основании этих данных Л.И. Колесниченко (1977) считает, что хромосомные числа у кувшинки не являются видовым признаком и отражают сложность популяционной структуры и историю развития этого рода.
В наблюдаемом в природе морфологическом многообразии кувшинок, можно выделить три составляющих: формы, обусловленные внутривидовым полиморфизмом, гибридные и экологические формы.
Из-за пространственной обособленности водоемов, заселенных кувшинкой, происходит ограничение скрещивания между особями пределами одной лишь заросли, что приводит к выделению местных форм с необычными комбинациями признаков (Комаров, 1937; Heslop-Harrison, 1955). В.Л. Комаров (1937) отмечает крайне слабую изученность этих форм и подчеркивает необходимость исследований живого материала, особенно формы и окраски рыльца, способных выявить еще неизвестные внутривидовые подразделения кувшинки. J. Heslop-Harrison (1955) отрицает возможность выделения таких форм в отдельные таксоны любого уровня.
Д.В. Дубына (1982) считает N. alba чрезвычайно морфологически изменчивым видом. В качестве наиболее изменчивого признака этот автор выделяет размер листовой пластинки. N. candida в Липецкой области характеризуется как весьма изменчивый вид с сильно варьирующим размером всех частей растения (Александрова и др., 1996), тогда как на Украине морфологические признаки этого вида колеблются в незначительной мере (Дубына, 1982). Наряду с типичной формой N. tetragona на протяжении всего ареала этого вида встречается форма с более узкими чашелистиками и лепестками, более многолепестным венчиком (10-15 лепестков) и более узким основанием чашечки. N. fennica Mela из Фенноскандии, является излишним названием, относящимся к типичной форме N. tetragona, что было признано и самим автором названия, поэтому это название не может быть использовано для узколепестной формы N. tetragona (Колесниченко, 1977). На Дальнем Востоке отмечена крупнолистная (22-27см х 13-16 см) и крупноцветковая (до 9 см в диаметре) форма N. tetragona, выделенная в отдельный вид N. wenzelii Maack (Колесниченко, 1977).
Существуют различные культурные формы кувшинок, которые впоследствии могут интродуцироваться: N. alba с красными чашелистиками (N. alba f. rosea C. Hartm) и с очень крупными и/или красно-пурпурными цветками (Uotilla, 2001); N. candida с кроваво-красной с фиолетовым оттенком (N. biradiata Sommer.) или желтой с красными точками на концах лучей (N. punctata Kar. et Kir.) окраской рыльца (Колесниченко, 1977; Крупкина, 2001), с розовыми (N. candida f. rosea Mela) или с красными цветками (N. candida f. erythropetala Mela и N. tetragona с изменяющими окраску цветками от белой через розовую до синевато-красной (N. tetragona f. colorata (Mela) Gurke) (Uotilla, 2001).
В местах симпатрического произрастания разных видов кувшинки наблюдается интенсивная межвидовая гибридизация, а также возвратные скрещивания с частым формированием гибридных популяций. Считается, что гибриды обладают промежуточной морфологией по сравнению с родительскими видами (Heslop-Harrison, 1955; Маевский, 1964; Куприянова, 1976; Цвелев, в печати), хотя по мнению Л.И. Колесниченко (1977), родительские признаки у гибридогенных форм существуют почти не изменяясь. R. Neuhausl и P. Tomsovic (1957, цит. по: Дубына, 1982) отрицают гибридное происхождение промежуточных по морфологии форм и указывают, что причиной существования этих форм в природе является широкая морфологическая изменчивость данных видов. К примеру, Д.В. Дубына (1982) отмечает существование на Украине большого числа переходных форм между N. alba и N. candida, но по неясным причинам относит все эти формы к N. alba. P. Uotilla (2001) вообще считает, что определение видовой принадлежности отдельных растений в гибридных популяциях становится невозможным.
Естественные гибриды N. alba x N. candida = N. x borealis Camus, возникшие в конце плейстоцена, имеют промежуточные признаки строения листьев и цветков (Heslop-Harrison, 1955; Маевский, 1964; Куприянова, 1976; Цвелев, в печати). По мнению В.Г. Папченкова N. x borealis характеризуется неясно четырехугольным основанием чашечки и яйцевидными очертаниями закрытого цветка и на территории Европейской России N. x borealis очень разнообразна. Она представлена как типичной, наиболее распространенной формой, у которой в равной степени сочетаются признаки родительских видов (N. x borealis var. intermedia), так и изредка встречаемыми N. x borealis var. percandida K. Valle (часто определялась в Средней и Северной России как N. candida) и N. x borealis var. peralba K. Valle (часто определялась в средней полосе как N. alba), у которых преобладают признаки либо одного, либо другого вида (Папченков, 2003).
Изредка, как правило, в местах совместного произрастания родительских видов, встречается гибрид между N. candida и N. tetragona – N. x sundvickii Hiit., характеризующийся более крупными по сравнению с N. tetragona размерами цветков (5-8 см в диаметре) и округло-эллиптических листьев, обратно-ланцетными нитями внутренних тычинок и сильно вдавленным красным или прурпурным рыльцем (Tutin, Webb, 1993). По мнению В.Г. Папченкова (2003), N. tetragona в Средней России отсутствует, а все сборы этого вида относятся к N. x sundvickii, хотя В.Л. Комаров (1937) указывал Карело-Лапландию как единственную возможную область произрастания N. x sundvickii в России.
Гибриды N. alba x N. tetragona характеризуются округлым основанием чашечки цветка, пурпурной окраской малолучевого сильно вдавленного рыльца с коническим центральным отростком цветоложа и расширенными нитями внутренних тычинок. Диаметр цветка такого гибрида составляет 5-6 см в диаметре, листовые пластинки эллиптические, 10-15 х 5-12 см (Комаров, 1937). Интересно, что иногда такие гибриды встречаются в отсутствие N. tetragona (Uotilla, 2001).
В мелких озерах и старицах, расположенных в лесной местности, в кислых олиготрофных водах с неблагоприятными гидрологическими (особенно температурными) и эдафическими условиями можно встретить кувшинки с мелкими цветами и листьями, меньшим числом лучей рыльца и более темной его окраской. В больших озерах мезотрофного типа, в старицах вдоль больших рек встречаются растения с более крупными цветками и листьями, большим числом лучей рыльца и более светлой его окраской (Heslop-Harrison, 1955, Куприянова, 1976; Колесниченко, 1977).
Опыты с искусственным изменением температуры и уровня воды показали, что размеры у кувшинки изменяются в зависимости от гидрологических условий, а не являются наследственными признаками (Дубына, 1982). Несмотря на кажущуюся ясной экологическую обусловленность существования мелко- и крупноцветковых форм, некоторыми авторами эти формы описываются как подвиды или даже особые виды: N. minoriflora (Borb.) Wissjul. или N. alba var. minoriflora Aschers. (Комаров, 1937; Куприянова, 1976; Колесниченко, 1977; Папченков, 2003), N. occidentalis (Ostenf.) Moss или N. alba subsp. occidentalis (Ostenf.) Hyl. (Heslop-Harrison, 1955), N. candida var. minor Wainio (Комаров, 1937; Колесниченко, 1977; Цвелев, в печати), тогда как существенных отличий в диагностических признаках, имеющих видовое значение, у этих форм обнаружено не было (Колесниченко, 1977).
В обсыхающих водоемах и на отмелях встречается мелколистная и мелкоцветковая форма N. alba с укороченными черешками -- N. alba f. terrestris Clavand. Такие растения после пересадки в более глубокий водоем приобретают типичную для N. alba морфологию (Heslop-Harrison, 1955).
Для N. alba была найдена зависимость размера отдельных частей растения от его возраста (Дубына, 1976). Вероятно, подобная зависимость существует и для других видов кувшинки.
Значения основных диагностических признаков для N. alba, N. candida и N. tetragona, содержащиеся в упомянутых выше определителях и обработках, обобщены в табл. 1.
|
признак |
N. alba |
N. candida |
N. tetragona |
|
форма основания чашечки |
округлая |
округленно-четырехгранная |
четырехугольная с выступающими ребрами |
|
форма нитей внутренних тычинок |
линейная |
ланцетная |
овальная |
|
скульптура экзины пыльцевых зерен |
шиповатая с длинными ворсинковидными и короткими столбовидными выростами |
бугорчатая |
мелкозернистая |
|
число лучей рыльца |
(7) 8-20 (23) |
6-14 (20) |
(4) 5-10 (16) |
|
форма рыльца |
почти плоское (слабо вогнутое) |
сильно вдавленное |
сильно вдавленное |
|
цвет рыльца |
желтое |
желтое, оранжевое, красное |
желтое, красное, пурпурное |
|
форма центрального отростка рыльца |
короткий полушаровидный |
длинный конический |
длинный конический |
|
диаметр цветка (см) |
(3) 5-15 (20) |
(3) 5-11 (16) |
3 (и меньше) –6 (10) |
|
внешний вид завязи |
не суживается к рыльцу и до верхушки покрыта рубцами от опавших тычинок |
суживается к рыльцу и не покрыта до верхушки рубцами от опавших тычинок |
|
|
форма бутона |
продолговато-яйцевидный с тупой верхушкой |
продолговато-яйцевидный с острой верхушкой |
четырехгранная пирамида |
|
форма листовой пластинки |
широко-эллиптическая, или округло-овальная, или округлая |
округло-эллиптическая, или округлая, или округло-овальная |
эллиптическая, или овальная, или округло-овальная |
|
форма крайних главных жилок листа |
почти прямая |
изогнута по всей длине |
изогнута только в первой трети |
|
длина листа (см) |
(10) 15-30 (35) |
(6) 12-26 (30) |
(4) 5-9 (20) |
|
ширина листа (см) |
(8) 14-27 (35) |
(8) 12-24 (30) |
(3) 4-10 (16) |
Во всех проанализированных источниках указываются сходные значения качественных диагностических признаков, четко разграничивающие в теории эти три вида кувшинки. Исключением является такой трудно формализуемый признак, как форма листовой пластинки, упоминаемый тем не менее в нескольких определителях (Маевский, 1976; Крупкина, 2001; Uotilla, 2001; Цвелев, в печати). Для характеристики таких трудно формализуемых качественных признаков, как форма листовой пластинки и формы ее крайних главных жилок, Л.А. Колесниченко (1977) и Д.В. Дубына (1982) используют коэффициент округлости листовой пластинки (отношение площади листовой пластинки к площади круга, описанного вокруг нее) и коэффициент кривизны крайних жилок (отношение длины перпендикуляра, восстановленного от линии, соединяющей место прикрепления черешка и край выреза листовой пластинки, к крайней жилке к длине этой линии). Очевидно, что такие коэффициенты чрезвычайно затруднительно использовать на практике, к тому же их значения для разных видов кувшинки различаются несущественно, что ставит под сомнение ценность этих признаков как диагностических. Различия в длине чашелистиков и наружных лепестков у разных видов кувшинки незначительны (Колесниченко, 1977; Цвелев, в печати), что также затрудняет использование этого признака в качестве диагностического. Приводимые в большинстве работ типичные значения количественных признаков для разных видов имеют широкий размах, заметно варьируют в зависимости от источника и сильно перекрываются между собой . Вероятно, причиной такой ситуации служит сильная вариабельность размеров кувшинки в зависимости от экологических условий и других факторов (Колесниченко, 1977; Дубына, 1982 и др.). Большая лабильность размеров листовой пластинки и затруднения в использовании формы листовой пластинки и формы ее крайних главных жилок в качестве диагностических признаков, вероятно, привели В.Э Скворцова (2000) к выводу о невозможности разграничения видов кувшинки по вегетативным признакам.
Многие основные диагностические признаки (цвет и форма рыльца, форма основания чашечки и т.д.) в роде Nymphaea меняются или исчезают даже при очень аккуратной гербаризации. Надо отметить, что в связи со спецификой объекта качественная гербаризация кувшинок затруднена и требует больших временных затрат при сушке (Лисицына, 2003). По этой причине чрезвычайно важно исследовать морфологию кувшинок на живом материале (Uotilla, 2001).
Для диагностики видов рода Nymphaea часто используют признаки пыльцевых зерен. Представители этого рода обладают своеобразным типом пыльцы, несвойственным другим группам покрытосеменных растений. Пыльцевое зерно Nymphaea состоит из двух полушарий, соединенных тонкой круговой мембраной. Дистальную отделенную мембраной часть называют крышечкой или оперкулумом, отчасти из-за того, что она отрывается при ацетолизной обработке или фоссилизации (Куприянова, 1976; Мейер-Меликян, Диамандопулу, 1996).
В экзине пыльцевых зерен покрытосеменных растений существуют борозды, поры и другие тонкие места, а иногда и сквозные отверстия, достигающие интины, которые объединяются под общим названием апертуры (Поддубная-Арнольди, 1976). Вопрос о том, к какому типу следует относить апертуру пыльцевого зерна Nymphaea активно обсуждался в литературе. M. Jones, G. Clarke (1981, цит. по Мейер-Меликян, Диамандопулу, 1996) и другие авторы предлагают считать апертуру кувшинок большой порой, расположенной на дистальном полюсе и почти полностью покрытой оперкулумом (однопоровые пыльцевые зерна). Некоторые исследователи (Снигеревская, 1955; Мейер, 1966 и др., цит. по Мейер-Меликян, Диамандопулу, 1996) считают бороздой лишь участок апертуры, не покрытый оперкулумом (зонально-бороздные пыльцевые зерна). Наконец, еще одна группа исследователей (Агабабян, 1976, цит. по Мейер-Меликян, Диамандопулу, 1996; Куприянова, 1976 и др.) апертурой считают всю дистальную часть пыльцевого зерна под оперкулумом. Последняя точка зрения подтверждается и сравнительно недавними исследованиями ультратонкого строения пыльцевых зерен Nymphaea (Мейер-Меликян, Диамандопулу, 1996). Таким образом, пыльцевые зерна Nymphaea следует считать дистально-однобороздными.
Существует мнение, что N. candida и N. alba могут быть уверенно разграничены лишь по строению пыльцевых зерен (Куприянова, 1976; Uotilla, 2001). Л.А. Куприянова (1976) показала, что пыльцевые зерна видов Nymphaea европейской части СССР характеризуются высокой морфологической стабильностью и четко различаются в основном по характеру скульптуры экзины. К сожалению, эта работа выполнена на очень малой выборке образцов (1-3 на вид), на основании морфологии пыльцы N. minoriflora и N. fennica выделены в отдельные виды, что позволяет считать результаты исследования Л.А. Куприяновой предварительными и требующими дальнейшей проверки. На этом литература, посвященная изучению скульптуры экзины у различных видов Nymphaea, исчерпывается (кроме библиографически редких работ K.J. Valle, 1927 и E. Pertola, 1962, 1975 на финском языке), хотя этот признак упоминается во многих распространенных обработках и определителях в качестве диагностического (Комаров, 1937; Дубына, 1982; Цвелев, 2000; Uotilla, 2001 и др.).
В.А. Поддубная-Арнольди указывала, что хотя строение, величина и форма пыльцевых зерен являются систематическими признаками, эти признаки могут значительно варьировать даже внутри одного вида. К примеру, полиплоидные формы имеют значительно более крупную пыльцу, чем диплоидные формы того же вида (Поддубная-Арнольди, 1976). В одной популяции N. candida пыльца из крупных и мелких цветков была фертильна, а из цветков среднего размера более или менее стерильна (Uotilla, 2001). У N. tetragona скульптура экзины пыльцевых зерен изменяется от мелкозернистой до крупнобугорчатой (Колесниченко, 1977).
Межвидовые гибриды кувшинки характеризуются пониженной фертильностью и разнообразной морфологией пыльцевых зерен (Комаров, 1937; Heslop-Harrison, 1955; Маевский, 1964; Куприянова, 1976; Цвелев, в печати). К примеру, пыльцевые зерна N. x borealis неоднородны в пределах одного цветка: они могут быть округлыми и непрозрачные или продолговатыми и прозрачными, иногда грушевидной формы, со скульптурой или полностью либо отчасти гладкие (Папченков, 2003), а для гибридов N. alba x N. candida характерна неясно шиповатая скульптура экзины пыльцевых зерен (Uotilla, 2001).
Из всего вышесказанного ясно, что существуют значительные трудности в разграничении видов кувшинки в европейской части России. Целью настоящей работы было как можно более подробное исследование морфологической изменчивости N. alba s.l. в европейской части России на живом материале.
В связи с этим перед нами были поставлены следующие задачи:
Популяция как элементарная эволюционная структура выделяется на основании географической и репродуктивной изоляции от других подобных структур (Тимофеев-Ресовский и др., 1977). Популяцией кувшинки можно назвать все растения кувшинки из одного водоема или водотока; за метапопуляцию мы принимали часть принадлежащих к данной популяции растений, четко обособленных пространственно. Таким образом, разными метапопуляциями мы считали группы кувшинок, произрастающих в одной реке на расстоянии 0,5-3 км друг от друга или в разных плесах одного обширного озера. Одним растением мы считали обособленную группу листьев и цветков.
Мы исследовали 42 метапопуляции кувшинки из 39 водоемов и водотоков следующих регионов европейской части России: Московская обл. (Волоколамский, Можайский и Одинцовский районы), Тверская обл. (Вышневолоцкий и Удомельский районы), Челябинская обл. (Кизильский район), Липецкая обл. (Грязинский район), республика Карелия (Лоухский район) и Мурманская обл. (Кандалакшский район). Мы старались включить в программу исследований те популяции кувшинки, о таксономической принадлежности которых были ранее сделаны заключения специалистами по этой группе (В.Г. Папченков, Л.И. Лисицына – ИБВВ АН РАН). Озерам, не имеющим названий на картах Генштаба масштаба 1: 50 000, были присвоены условные названия, указанные далее в тексте в кавычках (табл. 3). Мы отмечали основные характеристики водоемов и водотоков, в которых произрастали исследованные метапопуляции кувшинки (размер, характер зарастания, другие особенности).
|
номер метапопуляции |
название водоема или водотока |
индекс сапробности |
класс чистоты воды по: Сладечек, 1967 |
доля фертильных пыльцевых зерен (%) |
|
101 |
оз. Заверховье (Тверская обл.) |
1,7 |
3 |
100 |
|
102 |
оз. "Матрас" (Тверская обл.) |
1,4 |
2 |
100 |
|
103 |
оз. Клин (Тверская обл.) |
1,24 |
2 |
… |
|
104 |
оз. Боровно (Тверская обл.) |
1,4 |
2 |
… |
|
105 |
оз. Беленькое (Тверская обл.) |
1,4 |
2 |
100 |
|
106 |
оз. Головец (Тверская обл.) |
1,87 |
3 |
95 |
|
107 |
оз. Перхово (Тверская обл.) |
1,4 |
2 |
… |
|
108 |
вост. бер. оз. Молдино (Тверская обл.) |
1,6 |
3 |
… |
|
109 |
сев.-вост. берег оз. Молдино |
1,6 |
3 |
… |
|
110 |
центральная часть оз. Молдино |
… |
… |
99 |
|
111 |
оз. Рогозино (Тверская обл.) |
… |
… |
33 |
|
112 |
оз. Туришино (Тверская обл.) |
… |
… |
99 |
|
113 |
оз. Сорока (Тверская обл.) |
2 |
3 |
5 |
|
114 |
оз. Волково (Тверская обл.) |
1,5 |
3 |
97 |
|
115 |
оз. Писошно (Тверская обл.) |
1,4 |
2 |
95 |
|
116 |
оз. Почаево (Тверская обл.) |
1,5 |
3 |
63 |
|
117 |
пруд Стерляжий (Московская обл.) |
1,8 |
3 |
77; 87 |
|
118 |
озеро “Сима” (Московская обл.) |
0,95 |
2 |
… |
|
119 |
“пруд у турбазы” (Московская обл.) |
1,4 |
2 |
95 |
|
120 |
оз. "Индиго" (респ. Карелия) |
0,68 |
2 |
… |
|
121 |
р. Караганка (Челябинская обл.) |
1,9 |
3 |
93 |
|
127 |
оз. Моховое (Липецкая обл) |
… |
… |
91 |
|
128 |
р. Ресса выше по теч. (Калужская обл.) |
1,5 |
3 |
100 |
|
129 |
р. Ресса ниже по теч. (Калужская обл.) |
1,7 |
3 |
100 |
|
130 |
оз. "Тихое" (респ. Карелия) |
0,8 |
2 |
80 |
|
131 |
оз. "Гамарбия" (респ. Карелия) |
0,75 |
2 |
… |
|
132 |
мочажина у оз. Гремяха (Карелия) |
… |
… |
89 |
|
133 |
оз. Сенное (респ. Карелия) |
0,85 |
2 |
… |
|
134 |
оз. "Эврика" (респ. Карелия) |
0,7 |
2 |
100 |
|
135 |
оз. "Спелое" (респ. Карелия) |
0,7 |
2 |
93 |
|
136 |
оз. "Х" (респ. Карелия, Лоухский р-н) |
0,65 |
2 |
38 |
|
137 |
оз. "И" (респ. Карелия, Лоухский р-н) |
0,75 |
2 |
98; 88 |
|
138 |
оз. "Ла" (респ. Карелия, Лоухский р-н) |
0,78 |
2 |
94 |
Примечание: через точку с запятой указана доля фертильных зерен у разных цветков из одной метапопуляции
Мы гербаризировали измеренные цветок и лист от одного растения из каждой популяции и закладывали несколько тычинок этого цветка в силикагель для того, чтобы в последствии иметь возможность экстрагировать ДНК из собранных образцов.
Если число растений в метапопуляции не превышало 15, были исследованы все ее растения; если метапопуляция была более многочисленной, то исследовалось не менее 15 растений из нее. Таким образом, число исследованных растений в каждой метапопуляции колебалось от одного до 15 и в среднем было равно пяти. Всего исследовано 239 растений.
У каждого исследованного растения в полевых условиях мы регистрировали 9 качественных (форму бутона, цвет и форму рыльцевого диска, форму центрального отростка цветка, форму крайних жилок листа, форму нитей внутренних тычинок, форму основания чашечки, покрыта ли завязь до верхушки рубцами от тычинок, суживается ли завязь к рыльцу) и 6 количественных морфологических признаков (число лучей рыльца, длину, ширину и положение наибольшей ширины листовой пластинки, длину наружного лепестка и длину круга, образованного наружными тычинками), приводимых как диагностические в большинстве определителей и обработок (табл. 1).
Количество растворенной в воде органики является одним из немногих интегральных показателей состояния любых водных объектов, применяемых для оценки большинства факторов внешнего воздействия. Этот показатель отражает количество органических веществ, возникших в процессе жизнедеятельности организмов на всех трофических уровнях, а также внесенных с водосбора в результате природных и антропогенных процессов. Наиболее перспективным объектом для оценки состояния вод и экосистем, по мнению С.С. Бариновой (1992), являются водоросли – первичное и очень информативное звено трофических цепей. Водоросли встречаются практически во всех водоемах, поэтому оценка состояния водных экосистем, основанная на биоиндикации с помощью водорослей, позволяет сравнивать водные объекты разного типа и расположенные в разных регионах. При изменении содержания органических веществ в воде изменяется видовой состав индикаторных видов водорослей и их обилие.
Методы биоиндикации разрабатываются с начала 20 века. Наиболее распространенным методом в странах Восточной Европы и СНГ является метод Пантле-Бука, модифицированный В. Сладечеком (1967). Этот метод основан на понятии сапробности – способности организмов выживать в загрязненной органикой среде. Виды-индикаторы сапробности имеют в этой системе свой вес, выраженный в виде индекса сапробности S числом от 0 до 4. Индекс сапробности – это численное выражение способности сообщества гидробионтов выдерживать определенный уровень органического загрязнения. В системе Пантле-Бука и Сладечека индекс сапробности вычисляется по формуле: S= sum[s] x h/ sum[h], где s – сапробное значение организма-сапробионта в пробе, h – частота встречаемости сапробионта в пробе. Степень загрязнения в месте отбора пробы водорослей определяется по набору видов-индикаторов и по их обилию в пробе через индекс сапробности (Сладечек, 1967).
Для того чтобы оценить степень сапробности исследованных нами водоемов и водотоков, мы отбирали гидробиологическую пробу для каждого из них. При отборе пробы в пенициллиновый пузырек мы помещали немного разнообразного донного материала (ил, песок, грунт, камни, обрастания, растительные остатки), доливали пузырек примерно на 2/3 его объема водой из исследуемого водоема, добавляли 4-6 капель 40% формалина для консервации и герметично закрывали пузырек с указанием на нем номера метапопуляции согласно морфометрическому бланку. Дальнейшая обработка пробы велась на каф. микологии и альгологии биологического факультета МГУ под руководством и при участии О.В. Анисимовой.
Несколько капель пробы наносили на покровное стекло, смешивали с несколькими каплями 33% перекиси водорода и нагревали на асбестовом столике до полного выгорания органики, добавляя перекись водорода по мере ее испарения. Далее на предметное стекло помещали небольшое количество твердого канадского бальзама, расплавляли его на горелке и накрывали каплю канадского бальзама, образовавшуюся на предметном стекле, покровным стеклом с прокаленной пробой. Такая обработка позволяет получить препарат, на котором видны практически одни кремнеземные створками диатомовых водорослей, что существенно облегчает их определение. Расчет индекса сапробности производился по данным о видовом составе и по частоте встречаемости водорослей-индикаторов в пробе по методу Пантле-Бука в модификации В. Сладечека (1967).
В настоящее время появился существенно новый подход к изучению разнообразия форм объектов как таковых, исключая размерный фактор – геометрическая морфометрия (Bookstein, 1991; Adams et al., 2002). Две группы методов геометрической морфометрии распространены особенно широко: анализ Фурье для кривых, представляющих собой исследуемый контур, и методы, основанные на постановке меток на исследуемом контуре, такие как метод тонких пластин (TPS) (Adams et al., 2002). Первая группа методов позволяет построить описывающие форму объекта кривые и вычислить коэффициенты Фурье для этих кривых. Эти коэффициенты можно обрабатывать методами многомерного анализа данных. Вторая группа методов основана на постановке на контуре точек, отмечающих его ключевые места (так называемых “меток”). Эти метки должны быть гомологичны хотя бы геометрически, так как методы, основанные на метках, оперируют только с их координатами, и различия между объектами должны быть сравнимыми. В случае, когда строгая фиксация меток не представляется возможной из-за отсутствия четких “привязок”, допустимо использовать так называемые “полуметки”. Они расставляются равномерно в соответствии с заданным алгоритмом вдоль контура, число полуметок одинаково на всех сравниваемых контурах, причем первая и последняя из них обязательно фиксируются в строго определенных точках (Павлинов, 2001). TPS вычисляет степень деформации, необходимую для трансформации контура, образованного метками на одном образце, в такой же контур другого образца (прокрустова трансформация: Rolf, Slice, 1990). Многомерные методы анализа данных могут быть применены и к результатам TPS, так как проекция каждой формы на плоскость двух первых главных компонент, вычисленных из матрицы прокрустовых трансформаций, может играть роль обычной матрицы данных (Adams et al., 2002). Более подробный обзор методов геометрической морфометрии и ее математического аппарата, а также обширную библиографию по этой теме можно найти в статье И.Я. Павлинова и Н.Г. Микешиной (2002).
Подавляющее большинство публикаций, связанных так или иначе с методами геометрической морфометрии, выходит на английском языке, причем применению этого подхода для анализы формы ботанических объектов посвящено лишь несколько статей (например, Ray, 1992; Kores et al., 1993; Jensen et al., 2002). Русскоязычная литература по этой тематике невелика и посвящена в основном изучению зоологических объектов (Павлинов и др., 1994; Павлинов, 1999; Павлинов, 2000). Лишь в последнее время стали появляться немногочисленные русскоязычные статьи по применению методов геометрической морфометрии в ботанике (Волкова и др., в печати; Ильинский, Шипунов, в печати).
Контур гербаризируемого листа кувшинки сразу после его сбора мы обводили на бумаге. Для некоторых метапопуляций мы обводили контуры 10-15 листьев, взятых с одного и того же или разных растений, чтобы исследовать изменчивость формы листовой пластинки в пределах одного растения и внутри одной метапопуляции. Контуры удачно гербаризированных листьев (без явных признаков деформации их формы) мы также обводили на бумаге и измеряли, когда листья были полностью высушены. Мы также использовали контуры листовой пластинки N. candida и N. tetragona, заимствованные из “Flora Nordica” (Uotilla, 2001) и из “Illustrated Flora of Northern US and Canada” (Britton, Brown, 1913) в качестве “якорных образцов”.
Для характеристики формы листовой пластинки методами TPS (геометрическая морфометрия) мы использовали следующие подходы:
а) Расстановка шести биологически обусловленных меток на целой листовой пластинке (рис. 4а). К сожалению, такая расстановка меток, имея своим преимуществом истинную гомологичность соответствующих меток на разных листьях (тип I и тип II по Павлинов, 2001), не позволяет адекватно отразить форму листовой пластинки
Условные обозначения: пронумерованными кружками указано расположение меток
б) Расстановка разного числа меток вплоть до максимально допустимого из-за погрешностей их расстановки (26 меток на целой листовой пластинке и 14 меток на ее половине) большей частью по исключительно геометрическим соображениям (тип III по Павлинов, 2001) как на целой листовой пластинке, так и на ее половине (рис. 4б) с целью минимизации погрешностей при расстановке меток. Этот метод, позволяя адекватно отразить форму листовой пластинки, обладает одновременно большой погрешностью из-за большой зависимости месторасположения меток от положения контура листа в пространстве.
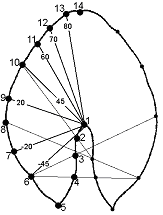
Условные обозначения: пронумерованными кружками указано расположение меток, числа на линиях обозначают угол (в градусах) между данной линией и горизонтальной линией
в) Анализ контура всей листовой пластинки (50 полуметок). При наличии перекрывания лопастей листа образуются два контура, что делает распознавание контура листовой пластинки программой TPSdig (Rohlf, 2001b) невозможным. Поэтому единственным условием применения этого подхода является удаление области перекрывания лопастей с изображения контура (4в), что ставит под сомнение адекватность этого способа анализа формы листовой пластинки.
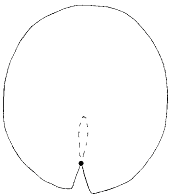
Условные обозначения: пунктиром показан удаленный контур перекрывающихся лопастей, кружочком показана начальная точка распознавания контура
г) Анализ контура половины листовой пластинки в целом (50 полуметок, рис. 4г). Недостатком этого подхода является включение в анализируемый контур искусственной прямой, разделяющей лист на две половины.
Условные обозначения: кружочком показана начальная точка распознавания контура
Использование эквивалентных меток типа III (Павлинов, 2001) позволяет сравнивать форму объектов как таковую, не получая представления о биологическом смысле обнаруженных различий (Kores et al., 1993). Вопрос об эффективности использования полуметок и об ограничениях, накладываемых на применение к ним методом геометрической морфометрии остается открытым. Тем не менее, в реальных ситуациях все-таки приходится использовать комбинацию из различных типов меток и полуметок (Павлинов, 2001).
Координаты точек снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig (Rohlf, 2001b). Координаты эталонной конфигурации, а также значения главных, относительных и частных трансформаций, характеризующие меру отличия образца от эталона, вычисляли при помощи программы tpsRelw, реализующей идею геометрической морфометрии в форме метода, аналогичного методу главных компонент (Rohlf, 2001a). Исходные координаты экземпляров нормировали относительно эталонной конфигурации при помощи прокрустова наложения, коэффициент шкалирования альфа принимали равным 0. Редактирование и конвертирование файлов данных мы производили при помощи вспомогательной программы tpsUtil (Rohlf, 2000).
Анализ данных методами геометрической морфометрии с использованием коэффициентов Фурье проводили при помощь программ EFAWin (Isaev, 1995) и MORPHEUS et al. (Slice, 1999).
Мы хотели сопоставить свои данные о размерах и скульптуре экзины пыльцевых зерен кувшинки с данными Л.А. Куприяновой (1976), полученными на ацетолизированном материале. Для этого пыльца из нескольких гербаризированных цветков была подвергнута ацетолизу по стандартной методике (Барыкина и др., 2000). Затем размеры пыльцевых зерен (большой и малый экваториальный диаметр) и характер скульптуры экзины ацетолизированной пыльцы были сопоставлены с аналогичными показателями для неацетолизированной пыльцы тех же цветков. Достоверные различия между размерами пыльцевых зерен одного и того же цветка до и после ацетолиза отсутствуют (малый экваториальный диаметр: t =1,8, p = 0,1; большой экваториальный диаметр: t = 0,8, p = 0,5), заметных изменений скульптуры экзины также не произошло. Поэтому все последующие исследования пыльцы под световым микроскопом мы проводили на интактной (не ацетолизированной) пыльце из гербаризированных цветков.
При изучении пыльцы под световым микроскопом мы приготовляли временный препарат, помещая пыльцу под покровное стекло в каплю глицерина, и просматривали препарат под микроскопом последовательно, измеряя все пыльцевые зерна, обнаруженные в подходящем положении (вид точно с полюса или точно с экватора). Морфология пыльцы исследовалась при помощи светового микроскопа МИКМЕД-1 на увеличении 1,5 x 15 x 40, все измерения проводили при помощи окуляр-микрометра (x 15) с точностью до 1 мкм. Всего было исследовано 368 пыльцевых зерен. Исследование скульптуры экзины пыльцы под световым микроскопом не позволяет уверенно отнести большинство просмотренных образцов к одному из типов, традиционно выделяемых в роде Nymphaea систематиками и полинологами (Куприянова, 1976; табл. 1). По этой причине мы осуществили дальнейшее исследование скульптуры экзины пыльцы при помощи сканирующего электронного микроскопа (далее СЭМ) в межфакультетской лаборатории электронной микроскопии. Для сопоставления с нашими образцами мы исследовали под СЭМ и пыльцу нескольких гербарных образцов из типичных местообитаний. Пыльцу прикрепляли к столику для СЭМ при помощи маникюрного лака, напыление проводили в ионнораспылительной установке IB-3 фирмы Eiko сплавом золота и палладия (примерная толщина напыления 25 нм). Микрографии получали при помощи сканирующего электронного микроскопа Camscan S-2 при ускоряющем напряжении 20 кВ в режиме SEI. Мы фотографировали первые увиденные в поле зрения 3-10 пыльцевых зерен, которые находились в нужном положении (вид с полюса или с экватора). Оцифровка изображения производилась при помощи компьютерной программы Scan Microcapture 2.20119 (разработчик А.В. Григорьев). Всего было сделано 272 микрофотографии.
Пыльца из гербаризированных цветков проверялась на фертильность путем ее окраски ацеткармином по стандартной методике без предварительной фиксации в реактиве Карнуа (Барыкина, 2000). Под световым микроскопом на увеличении 1,5 x 10 x 10 мы подсчитывали число окрашенных пыльцевых зерен, приходившееся на 100 случайно попавших в поле зрения зерен, вычисляя, таким образом, процент фертильных пыльцевых зерен в пробе (табл. 3).
С целью проверки полученных классификаций мы синтезировали четыре морфотипа (далее “якорных образца”), соответствующих “чистым” N. alba, N. candida, N. tetragona и N. x sundvickii на основе распространенных в литературе представлений о морфологии этих таксономических единиц.
Для выявлений различий и связей между переменными и построения классификаций использовались такие методы анализа данных, как дисперсионный анализ, непараметрический корреляционный анализ, непараметрический тест Уилкоксона и параметрический тест Стьюдента, а также анализ главных компонент. Тест на соответствие распределения значений данных нормальному был проведен при помощи вспомогательной программы, написанной А.Б. Шипуновым на основе теста Шапиро-Уилкса. Для всех вычислений использовалась компьютерная статистическая среда R (R Development Core Team, 2004).
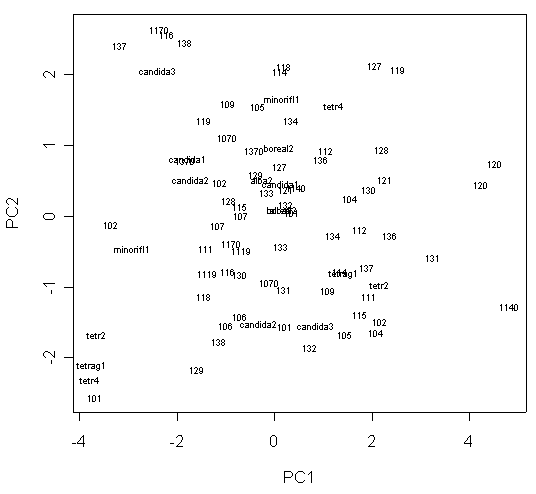
При анализе главных компонент матрицы морфометрических данных “якорный образец” N. candida располагается между “якорными образцами” N. alba и N. tetragona, а N. x sundvickii – между N. candida и N. tetragona, что полностью соответствует известным по литературе представлениям о морфологии этих таксономических единиц и свидетельствует об адекватности полученной классификации. На плоскости двух первых главных компонент растения образуют скорее единое облако вокруг “якорного образца” N. candida, чем отдельные группы. Исключение составляют лишь почти все растения из метапопуляции оз. “Гамарбия” и несколько растений из метапопуляции оз. “Тихое” респ. Карелия, тяготеющие к “якорному образцу” N. alba (рис. 5). Для этих растений характерна морфология, типичная для N. alba (табл. 1), за исключением округло-четырехугольной формы основания чашечки.
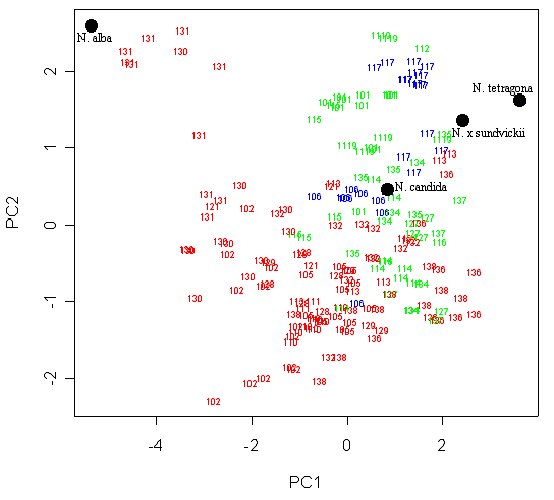
Условные обозначения: число обозначает одно растение, принадлежащее к метапопуляции с соответствующим номером (прил. 1), красным цветом обозначены растения с “мелкими” пыльцевыми зернами, зеленым – с “крупными”, синим – с пыльцевыми зернами “среднего размера” (см. Результаты). Черными точками обозначены “якорные образцы” (см. Материалы и методы, статистическая обработка данных).
Длина и ширина свежих листьев и тех же листьев, высушенных в гербарии, различается с высокой степенью достоверности (N=26, t-test: t=4,3; Df=45; p=0,0001 и t=4,8; Df=47; p=0,0002 соответственно). При гербаризации длина листьев уменьшается в 1,1-1,9 раз (среднее арифмитеческое +/- стандартное отклонение (SD) 1,3+/-0,2) – деформация листа в продольном направлении, ширина – в 1,0-1,9 раз (в среднем 1,4+/-0,2) – деформация листа в поперечном направлении. Коэффициент деформации листа (отношение степени деформации листа в продольном направлении к степени деформации листа в поперечном направлении) колеблется от 0,9 до 1,3 (в среднем 1+/-0,1). Таким образом, лист при высыхании уменьшается в размерах примерно в 1,5 раза. Степень деформации листа в продольном направлении достоверно не связана (r=0,3; p=0,1) со степенью деформации листа в поперечном направлении.
Результаты анализа главных компонент матрицы значений частных деформаций контуров листьев не позволяют выделить отдельных групп исследованных образцов по форме листьев. “Якорные образцы” N. candida и N. tetragona не различаются существенно между собой при всех способах расстановки меток и полуметок на контурах листовых пластинок с использованием TPS и при анализе формы листовых пластинок в целом с использованием коэффициентов Фурье. Самая удачная классификация (судя по расположению “якорных образцов”) получается при постановке 13 меток на половине листовой пластинке (рис. 6), но и она не позволяет выделять определенных групп на основании формы листовой пластинки. При всех подходах к анализу формы листовой пластинки кувшинки методами геометрической морфометрии разные листья одного растения или разных растений из одной метапопуляции не формировали единых групп, изменчивость внутри одной метапопуляции имела такой же масштаб, как и межпопуляционная изменчивость.
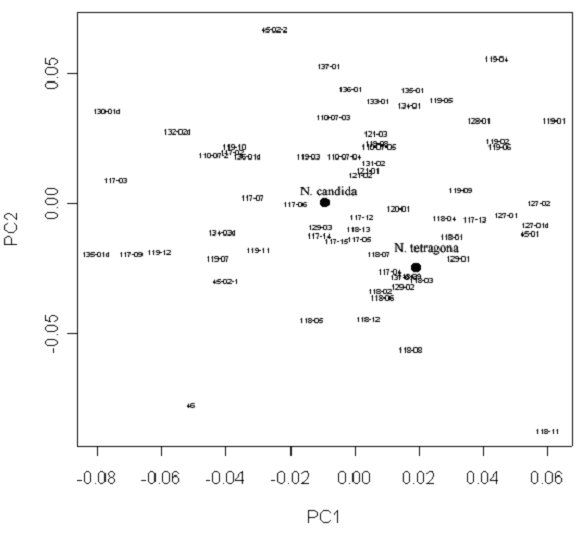
Условные обозначения: каждый лист обозначен как xx-yy, где xx – номер метапопуляции (согласно табл. 3), а yy – номер растения в этой метопопуляции согласно бланку. Черными точками обозначены “якорные образцы” (см. Материлы и методы, статистическая обработка данных).
Различия в степени сапробности (содержании органики) в исследованных водоемах и водотоках не велики. Значения индекса сапробности колеблются от 0,65 до 2,00, что соответствует второму и третьему классу чистоты воды (табл. 3). Большинство исследованных нами водоемов и водотоков олиго- или бетамезосапробные (индекс сапробности 1,4-2,0), а все карельские сплавинные озера и окно на болоте “Сима” Московской обл. относятся к олигоксено- или к ксенобетасапробным водоемам (индекс сапробности 0,65-1,00) по классификации В. Сладечека (1967).
Анализ главных компонент матрицы размерных характеристик кувшинки (число лучей рыльца, размеры цветка и листовой пластинки) не позволяет выделить отдельные метапопуляции с “мелкими” или “крупными” растениями. Мы не обнаружили статистически значимых корреляций размерных характеристик кувшинки со значением индекса сапробности или достоверных различий в размерах кувшинок, произрастающих в водоемах второго и третьего классов чистоты воды.
Размеры пыльцевых зерен (малый и большой экваториальный диаметр) у двух разных растений из одной метапопуляции достоверно отличаются в трех метапопуляциях из пяти исследованных (t-test, wilcox-test; p<0,05). Размеры пыльцевых зерен у растений из разных метапопуляций, принадлежащих одной популяции (р. Ресса Калужской области), также достоверно различаются (t-test; p<0,05).
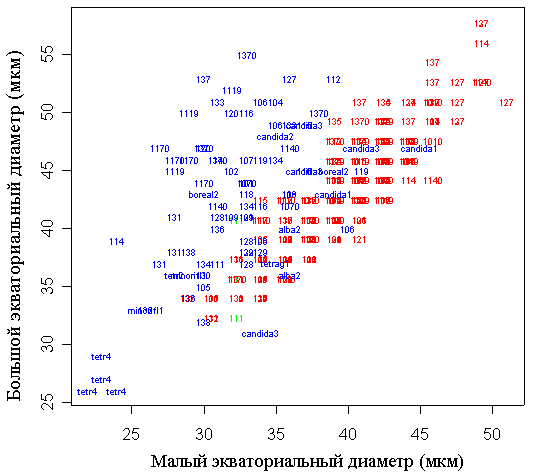
Значения малого и большого (рис. 7) экваториальных диаметров пыльцевых зерен имеют бимодальное распределение. Практически все исследованные метапопуляции по этому признаку четко разделяются на две группы с “мелкой пыльцой” (большой экваториальный диаметр 32-40 мкм, малый – 30-38 мкм) и с “крупной пыльцой” (44-52 мкм и 40-50 мкм соответственно). Две исследованные метапопуляции (оз. Головец Тверской области и пруд Стерляжий Московской обл.) имеют промежуточные размеры пыльцевых зерен (41-46 и 36-43 мкм соответственно). Значения отношения большого экваториального диаметра к малому имеют унимодальное распределение с максимумом на 1,03 и квартильным размахом 1,0-1,1.
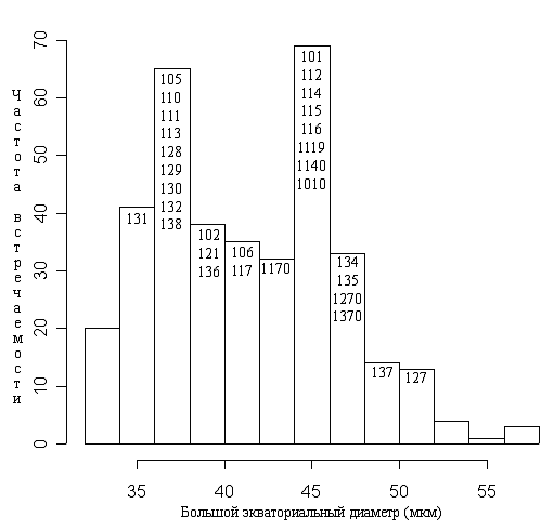
Условные обозначения: числа внутри столбцов диаграммы обозначают номера метапопуляций (согласно табл. 1), медианное значение максимального экваториального диаметра пыльцевых зерен которых совпадает с его значением для данного столбца. Номер метапопуляции с нулем на конце означает другой цветок этой метапопуляции.
Разделение метапопуляций на основании размера пыльцевых зерен соответствует их разделению по совокупности макроморфологических признаков (MANOVA: p<0,001), причем метапопуляции с пыльцевыми зернами промежуточного размера имеют морфологию близкую к морфологии метапопуляций с “крупными” пыльцевыми зернами (рис. 5). Однако выявить отдельные морфологические признаки, разграничивающие метапопуляции с разным размером пыльцы довольно затруднительно. Для этой цели удается использовать лишь квартильный размах значений цвета рыльца и числа его лучей. Для метапопуляций с “мелкими” пыльцевыми зернами характерно желтое рыльце с 11-13 лучами, для метапопуляций с “крупными” пыльцевыми зернами – желтое или оранжевое рыльце с 7-11 лучами, тогда как метапопуляциям с промежуточным размером пыльцевых зерен свойственно красное рыльце с 7-11 лучами.
Фертильность пыльцы из разных цветков, принадлежащих к одной метапопуляции, различалась не более чем на 10% (табл. 3). Растения из подавляющего большинства исследованных метапопуляций имеют высоко фертильную пыльцу (>75% фертильных пыльцевых зерен в пробе), для одной метапопуляции характерна пониженная фертильность пыльцы (63%): оз. Почаево Тверской области, две метапопуляции характеризуются низкой фертильностью пыльцы: оз. “Х” респ. Карелия (38%) и оз. Рогозино Тверской области (33%) и две – очень низкой (5%): оз. Сорока и оз. Клин Тверской области (табл. 3). Пыльцевые зерна в пыльниках растений из последней метапопуляции были чрезвычайно немногочисленны (10-20 на пыльник) и имели нетипичный внешний вид. Макроморфология растений из этой популяции также чрезвычайно своеобразна и сочетает в себе признаки N. alba (линейные нити внутренних тычинок, желтое и плоское рыльце), N. tetragona (практически четырехугольное основание чашечки) и N. candida (конический центральный отросток рыльца, главная жилка листа изогнута по всей длине).
Визуальный анализ полученных СЭМ-фотографий пыльцевых зерен показал, что скульптура экзины на общей поверхности пыльцевых зерен кувшинки неоднородна. Скульптура дистальной части пыльцевого зерна (крышечки), по нашему мнению, не демонстрирует дискретной изменчивости, представляя собой бугры разного размера, более крупные по краям крышечки. Скульптура экзины проксимальной части пыльцевых зерен чрезвычайно разнообразна. Далее мы будем понимать под скульптурой экзины пыльцевых зерен именно скульптуру их проксимальной части, как наиболее перспективную для выделения различных палинологических морфотипов. Можно выделить пять основных типов скульптуры экзины дистальной части пыльцевых зерен:
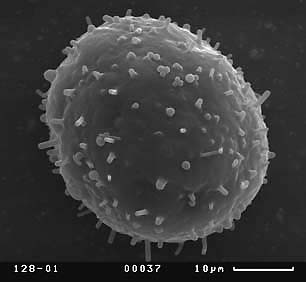



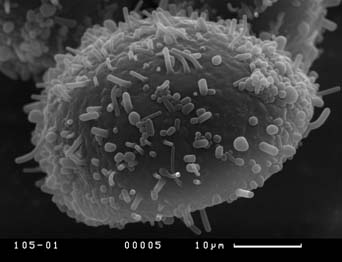
Скульптура экзины второго типа характерна для мелких пыльцевых зерен (35 x 37 мкм), тогда как скульптура экзины всех остальных типов характерна для пыльцевых зерен среднего и крупного размеров (рис. 7).
Надо отметить, что между этими выделяемыми нами типами скульптуры проксимальной части пыльцевых зерен существует непрерывный переход, что хорошо видно на рис. 10, где пыльцевые зерна не образуют сколь-нибудь четких групп при классификации их на основе размеров, формы и характера скульптуры экзины.
Условные обозначения: число обозначает пыльцевое зерно растения, принадлежащего к метапопуляции с соответствующим номером. Буквенные названия с числовым индексом обозначают пыльцу из гербарных образцов, см. Материлы и методы.
Нередко скульптура экзины существенно различается у разных растений из одной метапопуляции. Особенно интересной представляется нам метапопуляция кувшинки в пруду у турбазы Можайского р-на Московской обл. У южного берега пруда произрастает кувшинка с типичной для N. candida макроморфологией (табл. 1) и скульптурой экзины по типу 2. В центре пруда произрастает кувшинка некоторыми чертами макроморфологии, характерными для N. tetragona (изогнутые по всей длине главные жилки листа, сильно вогнутое рыльце, почти четерехгранное основание чашечки – см. табл. 1) и скульптурой экзины по типу 4. Скульптура экзины по типу 4 обнаружена нами еще в одной метапопуляции оз. Головец Тверской области, в которой макроморфология кувшинки отличалась от типичной для N. candida линейными нитями тычинок, изогнутыми по всей длине главными жилками листа и сильно вогнутым рыльцем (табл. 1).
Сканирующая электронная микроскопия, несомненно, позволяет более подробно изучить скульптуру экзины пыльцевых зерен. Обычной является ситуация, когда под световым микроскопом мы видим только палочковидные выросты, а на СЭМ фотографии наряду с палочковидными выростами становятся отчетливо заметны более мелкие бугорчатые или пузыревидные выросты.
Однако условия высокого вакуума, необходимого для СЭМ, приводят к заметной деформации пыльцевых зерен (рис. 9). Значения малого экваториального диаметра существенно уменьшаются при СЭМ (квартильный размах 30-36 мкм по сравнению с 37-46 мкм по данным промеров под световым микроскопом), эти различия высоко достоверны (t-test: p<<0,01). Значения большого экваториального диаметра у пыльцевых зерен после СЭМ достоверно не изменяются (t-test: p=0,56). Естественно, что при этом на СЭМ фотографиях пыльцевые зерна оказываются гораздо более вытянутыми, чем они есть на самом деле (квартильный размах отношения большого экваториального диаметра к малому 1,0-1,1 до СЭМ и 1,2-1,5 после СЭМ), эти различия высоко достоверны (t-test: p<<0,01).
Условные обозначения: числа и число-буквенные комбинации обозначают отдельные пыльцевые зерна, принадлежащие растениям из соответствующих метапопуляций (табл. 3) или гербарным образцам, цвет обозначает тип обработки пыльцевых зерен после гербаризации цветков: красный – отсутствие последующей обработки, зеленый – ацетолиз, синий – сканирующая электронная микроскопия
Наши исследования показали, что морфология кувшинки существенно изменяется при традиционных способах обработки живого материала. При гербаризации непредсказуемым образом искажаются размер и форма листьев, а также необратимо меняются важные диагностические признаки (например, цвет и форма рыльца). Этот факт справедливо отмечался ранее рядом исследователей (Комаров, 1937; Uotilla, 2001; Лисицына, 2003), хотя количественные исследования изменений макроморфологии кувшинки при гербаризации на конкретных образцах были впервые проделаны лишь в настоящей работе. В связи с этим нужно более критически относиться к систематическим обработкам рода, так как они в основном проводятся на основе гербарного материала (Комаров, 1937; Маевский, 1974; Лисицына и др., 1993; Губанов и др., 1995), что вызывает дополнительные трудности в диагностики видов кувшинки. По нашему мнению, исследования макроморфологии кувшинки должны проводиться на живом материале.
Размерные характеристики пыльцевых зерен существенно искажаются при обработке на СЭМ. В связи с этим промеры пыльцевых зерен кувшинки на СЭМ-микрографиях, сделанные J.B. Muntendam с соавторами (1996), нельзя считать адекватными. Ацетолизная обработка, с одной стороны, не изменяет существенно размерных характеристик пыльцевых зерен, а, с другой стороны, не облегчает заметно анализ скульптуры экзины, поэтому применение трудоемкой методики ацетолиза для изучения палиноморфологии кувшинки неоправданно. Надо отметить, что без применения СЭМ невозможно в полной мере исследовать скульптуру экзины пыльцевых зерен. Исследования морфологии пыльцевых зерен кувшинки на малых выборках, как это было проделано Л.А. Куприяновой (1976), некорректны из-за большой изменчивости палиноморфологии даже в пределах одной метапопуляции. Таким образом, для исследования палиноморфологии кувшинки необходимо использование как световой, так и электронной микроскопии на больших выборках пыльцевых зерен без их ацетолизной обработки.
В связи с тем, что наши данные не позволяют выделить дискретных морфотипов кувшинки на исследованной территории, нам сложно оценить диагностическую ценность отдельных макроморфологических признаков. Кроме того, такая оценка вряд ли необходима, особенно в свете мнения Л.И. Колесниченко (1977) о необходимость использования комплекса морфологических признаков для систематики рода Nymphaea из-за того, что по отдельным признакам европейские виды кувшинки сближаются друг с другом.
Нам все же хотелось бы отметить полную несостоятельность такого часто упоминаемого в определителях и обработках (Комаров, 1937; Маевский, 1976; Лисицына и др., 1993; Крупкина, 2001; Uotilla, 2001) диагностического признака, как форма листовой пластинки. В настоящей работе было впервые проведено количественное изучение формы листовой пластинки как таковой, без учета ее размерных характеристик, методами геометрической морфометрии. В результате этого исследования было показано, что форма листовой пластинки в пределах одного растения варьирует так же сильно, как и у растений, принадлежащим к разным популяциям. Кроме того, форма листовой пластинки заметно меняется при гербаризации, а ее словесное описание затруднено.
Достоверная зависимость размерных характеристик кувшинки от степени сапробности водоема нами не выявлена, что противоречит распространенному в литературе мнению (Комаров, 1937; Heslop-Harisson, 1955; Куприянова, 1976; Колесниченко, 1977; Дубына, 1982; Папченков, 2003). С одной стороны, можно предположить, что разброс значений степени сапробности исследованных нами водоемов был недостаточен для выявления подобной зависимости. С другой стороны, ранее специальных количественных исследований степени сапробности водоемов не проводили, а оценивали ее на основании общих соображений. Небольшие северные сплавинные озера традиционно относятся к олиготрофным в противоположность мезотрофным озерам средней полосы России (Куприянова, 1976; Колесниченко, 1977), в то время как наши количественные исследования не демонстрируют существенных различий в степени сапробности между исследованными озерами. Можно предположить, что существование мелких форм кувшинки не так четко обусловлено степенью сапробности водоема, как это считалось ранее.
Наши исследования морфологии кувшинки не подтвердили сведения о произрастании на исследованной территории N. alba, N. tetragona, N. x borealis и N. x sundvickii (Лисицына и др., 1993; Головина и др., 2003; Папченков, 2003; Лисицына, личное сообщ.). По нашим данным, многообразие морфотипов кувшинки на исследованной территории нельзя разделить на более или менее дискретные группы. Причиной этого, вероятно, является сложное сочетание двух противоположных процессов, интенсивно происходящих в роде Nymphaea на севере европейской части России: (1) активная межвидовая гибридизация и интрогрессия во время плейстоцена из-за наступления ледника (Muntendam et al., 1996) и в настоящее время вследствие снятия фенологической изоляции из-за нестабильных климатических условий (Ветчинникова, 2004) и (2) обособление локальных форм из-за изолированности водоемов и интенсивных хромосомных перестроек и генных мутаций (Комаров, 1937; Heslop-Harrison, 1955). Отдельные крайние морфотипы существующего морфологического континуума могут трактоваться как N. alba и N. tetragona, а образцы, занимающие по морфологии промежуточное положение между воображаемым центром континуума (типичная N. candida) и его крайними морфотипами -- как гибридные формы (N. alba x N. candida = N. x borealis и N. tetragona x N. candida = N. x sundvickii). Подобный подход доведен до своего логического завершения В.Г. Папченковым (2003). Необходимо отметить, что такая трактовка не соотвествует общепринятому определению вида (Тимофеев-Ресовский и др., 1977) и не может считаться справедливой. Недопустимость выделения видов на основании небольших отличий в морфологии особенно актуальна для растений, у которых вегетативное размножение преобладает над семенным (Elven et al., 2004), таких как кувшинка (Heslop-Harrison, 1955; Дубына, 1982).
Нашу точку зрения подтверждают и сведения о произрастании “гибридных форм” в отсутствие родителей (Колесниченко, 1977; Uotilla, 2001). Насколько нам известно, в этих исследованиях гибриды выделялись только по морфологии, и за “гибридные формы” легко могли быть приняты растения с нетипичными комбинациями морфологических признаков, не имеющие гибридного происхождения. Проведенные нами исследования фертильности пыльцы показали, что растения с нетипичной для чистых видов морфологией, как правило, имеют высоко фертильную пыльцу и вряд ли могут считаться гибридами, что подтверждает мнение R. Neuhausl и P. Tomsovic (1957, цит. по: Дубына, 1982).
Наши данные не демонстрируют приуроченности различных морфотипов к определенным географическим регионам. Поэтому выделение их даже в ранг подвида на данном этапе исследований не обоснованно (Elven et al., 2004). Для выяснения географической приуроченности различных морфотипов необходимо провести продолжить наши исследования в южной части Европейской России, где ожидается типичная N. alba, и в Сибири, где ожидается типичная N. tetragona (Комаров, 1937; Muntendam et al., 1996).
Четкое разделение исследованных метапопуляций кувшинки по размеру пыльцы, соответствующее их разделению лишь по совокупности макроморфологических признаков, но не по отдельным признакам, позволило нам сделать предположение о существовании двух хромосомных рас кувшинки на исследованной территории. Это согласуется с данными о скоррелированности размера пыльцевых зерен с уровнем плоидности (Поддубная-Арнольди, 1976) и о большом числе известных хромосомных чисел для европейских видов кувшинки (Heslop-Harisson, 1955; Дубына, 1982; Wiersema, 1988; Крупкина, 2001). В этом случае легко предположить, что растения с разным уровнем плоидности несколько отличаются друг от друга по комплексу макроморфологических признаков. Для проверки этой гипотезы в дальнейшем оценивать уровень плоидности исследуемых растений.
По результатам настоящей работы могут быть сделаны следующие выводы:
Для уточнения сложившейся ситуации с морфологическим разнообразием в роде кувшинка в Европейской России в следующем полевом сезоне необходимо расширить географию исследований до южной части Европейской России и Сибири, а также оценивать уровень плоидности исследуемых растений.
Мы искренне благодарны всем, кто оказывал нам всестороннее содействие при сборе материала в полевых условиях и при его дальнейшей обработке: участникам летних практик Московской Гимназии на Юго-Западе (№1543), проходивших под руководством С.М. Глаголева; зам. директора по НИР природно-исторического заповедника “Аркаим” Е. Чибилеву; зам. директора по НИР государственного природного Кандалакшского заповедника А.С. Корякину и доценту Липецкого университета К.И. Александровой. Мы также благодарны сотруднице каф. высших растений биологического факультета МГУ С.В. Полевовой за консультации в области палинологии, сотруднице каф. альгологии и микологии биологического факультета МГУ О.В. Анисимовой за помощь в обработке гидробиологических проб и консультации по определению степени сапробности водоемов, А.Б. Шипунову и С.Р. Майорову за конструктивное обсуждение работы, ведущему инженеру МЛЭМ А.Г. Богданову за компетентную и доброжелательную помощь на сеансах электронной микроскопии, К. Марквичевой за подготовку части иллюстраций и С. Назаровой за предоставление литературы.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |