В местах совместного произрастания Drosera rotundifolia L. и D. anglica Huds. встречается их гибрид D. ×obovata Mert. et Koch (Schnell, 1999). Существование различных морфологических переходов между D. anglica, D. ×obovata и D. rotundifolia (Wood, 1955; Schnell, 1999), а также разнообразие экологических форм этих таксонов затрудняют их разграничение (Волкова, 2002; Волкова и др., 2003). Основными определительными признаками являются метрические характеристики формы и размера листьев, а также их положения относительно субстрата (Раменская, Андреева, 1982; Кирпичников, 1964 и др.). В качестве дополнительного определительного признака C.E. Wood (1955) предложил использовать форму поперечного среза черешков. Однако эта идея не учитывается при составлении определителей (Раменская, Андреева, 1982; Кирпичников, 1964 и др.), вероятно, из-за трудностей формализации этого признака и его использования на практике.
В настоящее время появился существенно новый подход к изучению разнообразия форм объектов как таковых, исключая размерный фактор – геометрическая морфометрия. Подавляющее большинство публикаций, связанных так или иначе с методами геометрической морфометрии, выходит на английском языке, причем применению этого подхода для анализы формы ботанических объектов посвящено лишь несколько статей (например, Ray, 1992; Kores et al., 1993). Русскоязычная литература по этой тематике невелика и посвящена изучению зоологических объектов (Павлинов и др., 1994; Павлинов, 1999; Павлинов, 2000). Обзор методов геометрической морфометрии и ее математического аппарата, а также библиографию по этой теме можно найти в статье И.Я. Павлинова и Н.Г. Микешиной (2002). Вопрос о соответствии результатов, полученных методами геометрической морфометрии, сложившимся на основе традиционных методов анализа данных представлениям по-прежнему остается открытым (Павлинов, 2000). К примеру, методы геометрической морфометрии при исследовании формы жевательной поверхности зубов у полевок позволяют выявить иные элементы дискретности, чем методы классической морфометрии (Павлинов, 1999).
Целью настоящей работы является сопоставление классификации ботанических объектов методами геометрической морфометрии и классической морфометрии.
Сбор материала проводили в июне-августе 2002 и 2003 годов в Удомельском и Вышневолоцком районах Тверской области и Лоухском районе республики Карелия. Всего исследовали четыре популяции D. rotundifolia (одну в республике Карелия и три в Тверской области), одну популяцию D. ×obovata и одну популяцию D. anglica из Карелии (табл. 1).
Таблица 1. Основные характеристики исследованных популяций Drosera spp.
|
№ попу- |
вид |
место сбора |
местообитание |
дата сбора |
автор сбора |
число образцов для геометрич. морфометрии |
число образцов для классич. морфометрии |
|
1 |
D. rotundifolia |
Тверская обл., Удомельский р-н, пос. Еремково, оз. Гнилое, около завода |
на сплавине |
22.06.2003 |
И. Юфряков |
14 |
0 |
|
2 |
D. rotundifolia |
Тверская обл., Удомельский р-н, пос. Еремково, оз. Гнилое, вост. берег |
на сплавине |
22.06.2003 |
И. Юфряков |
15 |
0 |
|
3 |
D. rotundifolia |
Тверская обл., Вышнево-лоцкий р-н, зап. берег оз. Ольшево |
на сплавине |
02.07.2003 |
И. Юфряков |
14 |
25 |
|
4 |
D. ×obovata |
респ. Карелия, Лоухский р-н, 250 м к северу от дер. Нижняя Пулонга, сев. берег оз. Черное |
на сплавине |
13.08.2003 |
П. Волкова, И. Юфряков |
15 |
17 |
|
5 |
D. anglica |
респ. Карелия, Лоухский р-н, южнее усадьбы ББС МГУ, у нижней части “Ботанического сада” |
верховое болотце у ручья |
10.07.2002 |
П. Волкова |
17 |
13 |
|
6 |
D. rotundifolia |
респ. Карелия, Лоухский р-н, южнее усадьбы ББС МГУ, у нижней части “Ботанического сада” |
верховое болотце у ручья |
10.07.2002 |
П. Волкова |
15 |
28 |
|
7 |
D. rotundifolia |
США, северный берег южного полуострова оз. Мичиган |
1953 |
C.E. Wood (1955, стр. 110, рис. 25) |
1 |
0 |
|
|
8 |
D. linearis |
США, северный берег южного полуострова оз. Мичиган |
1953 |
C.E. Wood (1955, стр. 110, рис. 30) |
1 |
0 |
Для получения контуров поперечного среза черешка из каждой популяции выбирали 14-17 растений. Бритвой делали тонкий поперечный срез срединной части черешка наибольшего ловчего листа каждого растения. Качество среза контролировали под бинокуляром. Затем из среза изготовляли временный препарат, контур которого при помощи микроскопа (увеличение 56´) в комплекте с рисовальным аппаратом РА-6 переносили на лист бумаги. Также мы использовали контуры поперечного среза срединной части черешка D. rotundifolia и D. linearis (родительских форм D. anglica) из работы C.E. Wood (1955). Полученные контуры поперечных срезов вводили в компьютер через планшетный сканер.
Для четырех исследованных популяций были также сделаны промеры длины черешка наибольшего листа, длины и ширины листовой пластинки наибольшего листа, расстояния от ее основания до положения ее наибольшей ширины, а также отмечено положение наибольшего листа относительно субстрата. Для этих промеров выбирали 13-28 не использованных для получения контура поперечного среза черешка растений из каждой популяции. Таким образом, данные для классической и геометрической морфометрии собирались для популяции в целом, поскольку возможные различия между видами лучше всего заметны на популяционном уровне.
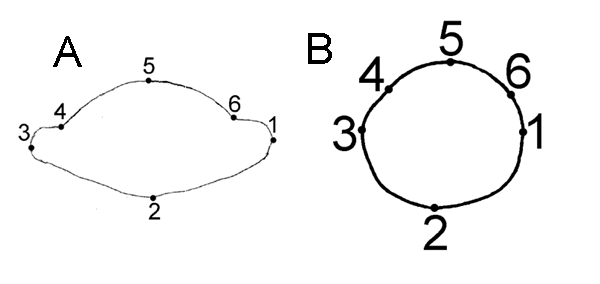
Для характеристики формы поперечного среза черешка использованы 6 меток, расположенных на его контуре. В нашем случае для фиксации меток использовались не только биологические, но и геометрические соображения: метки расставлялись в точках наибольшей кривизны контура (рис. 1), что позволяет отнести применяемые нами метки к типу II (Павлинов, Микешина, 2002). Использование таких геометрически гомологичных меток позволяет сравнивать форму объектов как таковую, не получая представления о биологическом смысле обнаруженных различий (Kores et al., 1993).
Рисунок 1. Форма поперечного среза срединной части черешка у Drosera rotundifolia и D. linearis: положение точек для снятия координат
1a D. rotundifolia
1b D. linearis (по Wood, 1955 с изменениями)
Координаты точек снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig (Rohlf, 2001b). Координаты эталонной конфигурации, а также значения главных, относительных и частных трансформаций, характеризующие меру отличия образца от эталона (Павлинов, Микешина, 2002) вычисляли при помощи программы tpsRelw, реализующей идею геометрической морфометрии в форме метода, аналогичного методу главных компонент (Rohlf, 2001a). Усредненные контуры поперечного среза срединной части черешка для каждого вида получали при помощи программы tpsSuper (Rohlf, 2003). Редактирование и конвертирование файлов данных мы производили при помощи вспомогательной программы tpsUtil (Rohlf, 2000).
Для анализа межпопуляционных различий удобно использовать не отдельные особи, а некие средние для каждой популяции образцы, обладающие характерными для большинства особей данной популяции значениями исследуемых признаков. Подобный подход не только делает анализ более наглядным, но и позволяет избавиться от «выбросов» (нетипичных образцов). В качестве конфигурации, адекватно представляющей популяцию для геометрической морфометрии, мы выбрали два варианта. Во-первых, искусственно сгенерированную усредненную конфигурацию (“consensus configuration”) и, во-вторых, такую реальную конфигурацию (назовем ее “центральной особью”, ЦО), которая в пространстве относительных деформаций располагается максимально близко к началу осей, то есть к эталонной конфигурации (Павлинов, Микешина, 2002).
Для классической морфометрии популяции были представлены данными, состоящими из медиан каждого признака. Многомерное шкалирование, анализ главных компонент и кластерный анализ (метод Уорда, манхеттеновская метрика) матриц значений частных деформаций и шкалированных результатов морфологических промеров были проделаны при помощи пакета программ R 1.7.1 (Venables et al., 2002). При многомерном анализе данных угол наклона листа относительно субстрата выражали в радианах.
В пространстве двух измерений многомерного шкалирования матрицы значений частных деформаций индивидуальные конфигурации образуют три “облака” в соответствии с видовой принадлежностью образцов, отдельно располагается D. linearis (рис. 2a). Наблюдаемая размытость “облаков” и наличие нескольких выбросов свидетельствуют, вероятно, о методических неточностях при получении контуров поперечных срезов. Между группами, образованными особями из популяций D. rotundifolia и D. anglica, располагается группа, образованная особями D. ×obovata. Интересно отметить, что конфигурации из популяций D. rotundifolia не разделяются в соответствии с их принадлежностью к определенной популяции и/или региону (рис. 2b).
Рисунок 2. Распределение исследованных растений в пространстве двух измерений многомерного шкалирования частных деформаций
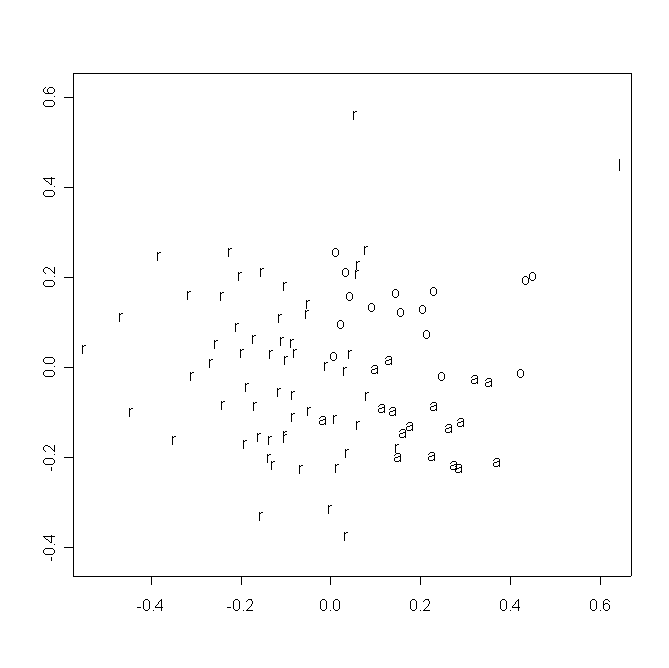
2a. Условные обозначения:
r Drosera rotundifolia
o D. ×obovata
a D. anglica
l D. linearis
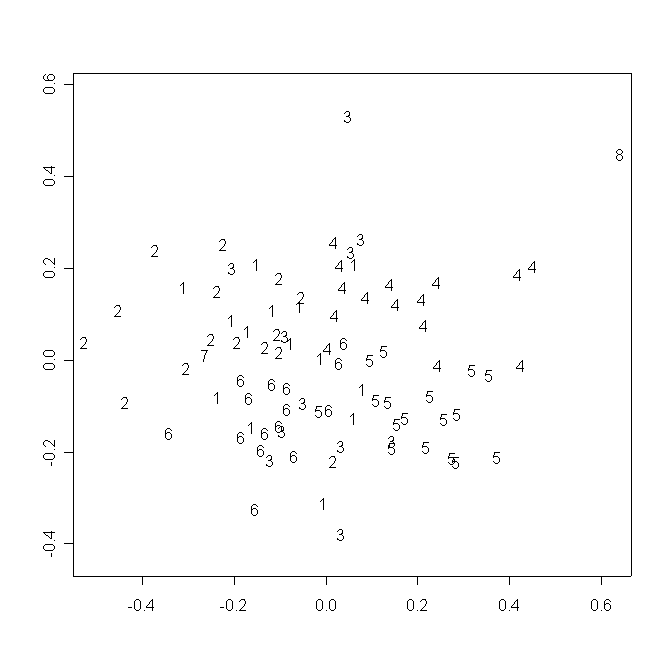
2б. Условные обозначения:
Одинаковыми номерами обозначены растения
из одной популяции. Номера популяций
соответствуют номерам в табл. 1.
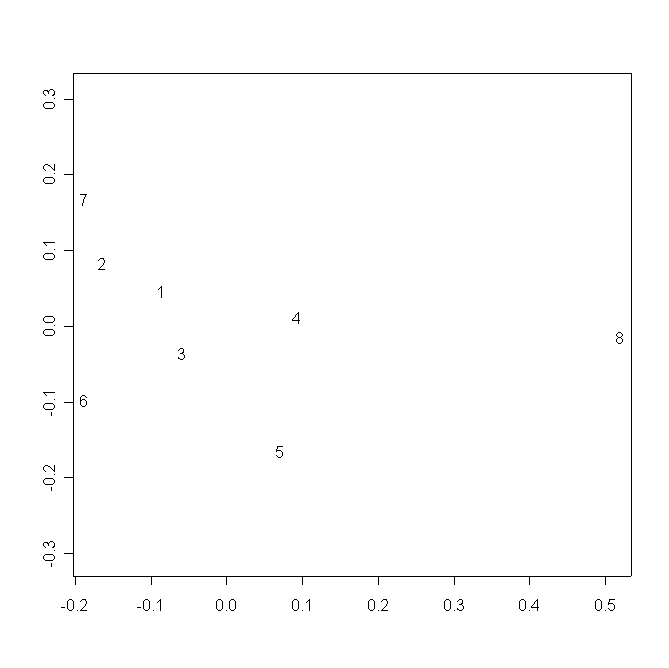
В аналогичном пространстве ЦО всех популяций D. rotundifolia образуют единую группу, четко дистанцированную от ЦО популяции D. anglica. Между группой популяций D. rotundifolia и ЦО D. anglica (ближе к первой), располагается ЦО популяции D. ×obovata. D. linearis располагается обособлено, ближе к ЦО D. anglica (рис. 3).
Рисунок 3. Распределение ЦО в пространстве двух измерений многомерного шкалирования частных деформаций
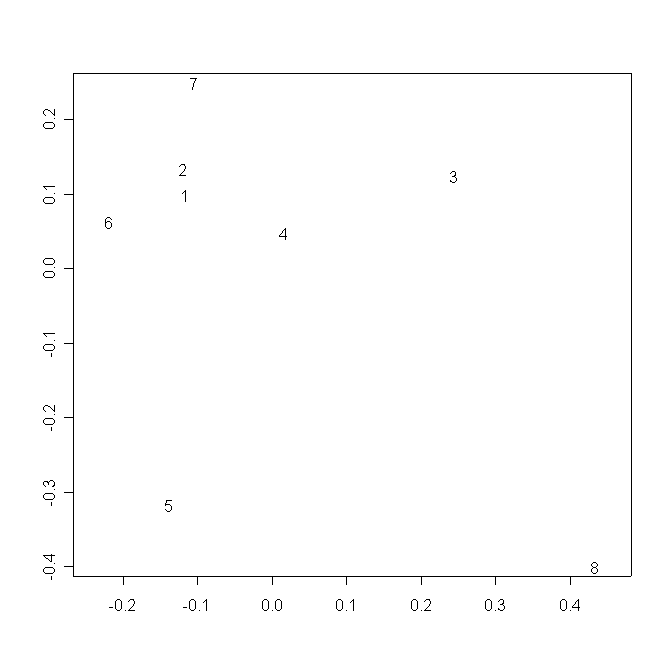
Условные обозначения:
См. условные обозначения к рис. 2б.
При подобном анализе для усредненных конфигураций наблюдается сходная картина (рис. 4).
Рисунок 4. Распределение усредненных конфигураций в пространстве двух измерений многомерного шкалирования частных деформаций
Условные обозначения:
См. условные обозначения к рис. 2б.
При кластерном анализе матрицы частных деформаций для ЦО выделяются два кластера. Один из них объединяет D. rotundifolia, произрастающую на оз. Ольшево (Тверская область), D. anglica и D.linearis; второй кластер включает D. rotundifolia из других популяций и D. ×obovata (рис. 5).
Рисунок 5. Кластерный анализ матрицы частных деформаций для ЦО
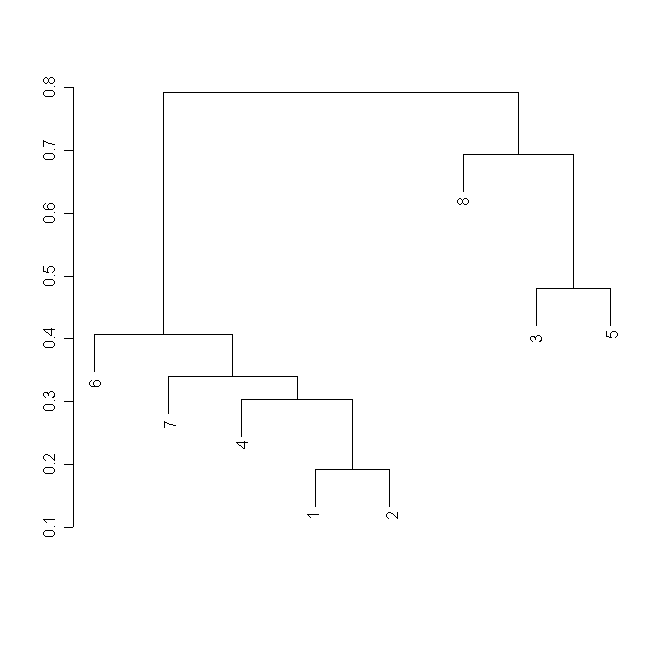
Условные обозначения:
См. условные обозначения к рис. 2б.
В кластерном анализе для усредненных конфигураций выделяются две группы: одна группа (I) представлена D. linearis, другая же разделяется на три подгруппы. Подгруппы II и III объединяют усредненные конфигурации всех исследованных популяций D. rotundifolia, тогда как подгруппа IV состоит из усредненных конфигураций популяций D. ×obovata и D. anglica (рис. 6).
Рисунок 6. Кластерный анализ матрицы частных деформаций для усредненных конфигураций
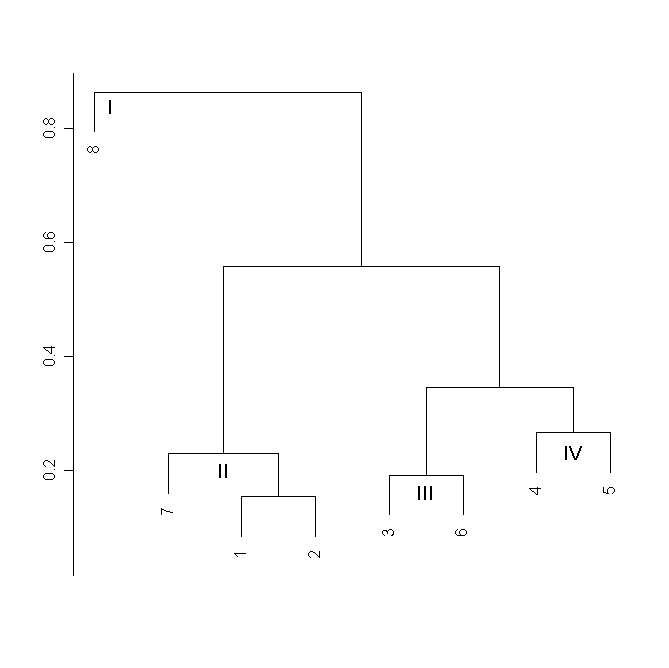
Условные обозначения:
См. условные обозначения к рис. 2б.
Распределение нагрузок на относительные деформации дает следующие результаты (табл. 2): первая деформация более всего связана с первой и шестой метками, а вторая деформация – с первой и второй метками (см. также рис. 1). Таким образом, наибольшим изменениям подвержена правая абаксиальная часть среза, что является довольно неожиданным результатом из-за билатеральной симметрии полученных контуров.
Таблица 2. Нагрузки на первую и вторую относительные деформации поперечного среза срединной части черешка исследованных видов Drosera spp.
|
номер метки |
нагрузки |
|||
|
RW1 |
RW2 |
|||
|
X |
Y |
X |
Y |
|
|
1 |
~0 |
15,3 |
5,4 |
~0 |
|
2 |
1,2 |
~0 |
~0 |
6,4 |
|
3 |
~0 |
9,8 |
-0,5 |
~0 |
|
4 |
10,9 |
~0 |
2,5 |
0,3 |
|
5 |
~0 |
8,8 |
0,1 |
4,4 |
|
6 |
13,4 |
~0 |
~0 |
1,5 |
В пространстве двух измерений многомерного шкалирования данных морфологических промеров особи образуют три “облака” в соответствии с видовой принадлежностью образцов. Между группами, образованными особями из популяций D. rotundifolia и D. anglica (ближе к первой группе), располагается D. ×obovata. Растения D. rotundifolia из республики Карелия и Тверской области располагаются в противоположных частях соответствующего “облака”, смешиваясь лишь в центральной его части (рис. 8).
Рисунок 8. Распределение исследованных растений в пространстве двух измерений многомерного шкалирования результатов морфологических промеров
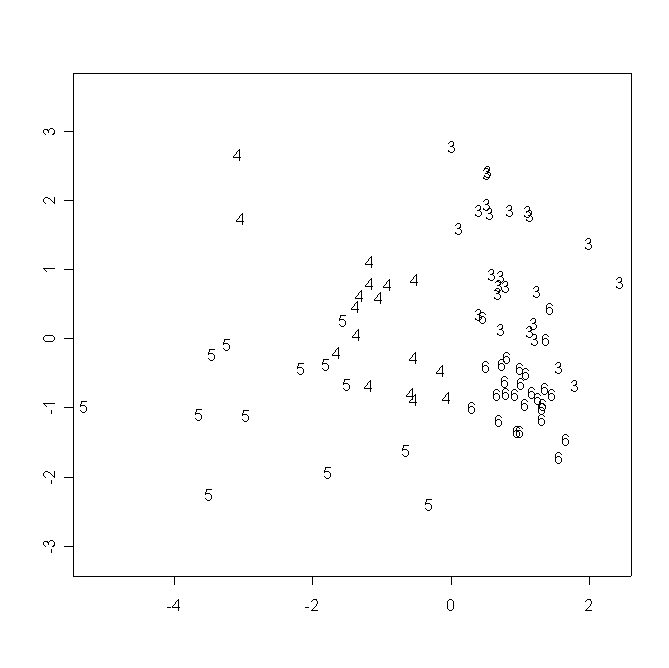
Условные обозначения:
См. условные обозначения к рис. 2б
Анализ нагрузок на оси главных компонент показал, что наиболее важными для классификации признаками являются (а) длина листовой пластинки и (б) расстояние от основания листовой пластинки до уровня ее наибольшей ширины (табл. 3).
Таблица 3. Нагрузки шкалированных значений линейных промеров на оси главных компонент
|
промеры |
нагрузки |
|
|
PC1 |
PC2 |
|
|
длина листовой пластинки |
0,64 |
~0 |
|
ширина листовой пластинки |
-0,32 |
-0, 65 |
|
расстояние от основания листовой пластинки до уровня ее наибольшей ширины |
0,64 |
~0 |
|
длина черешка |
0,26 |
-0,65 |
|
угол между черешком и субстратом (в радианах) |
0,15 |
-0, 38 |
В аналогичном пространстве популяции D. rotundifolia обособлены от D. anglica и D. ×obovata (рис. 9).
Рисунок 9. Распределение популяций в пространстве двух измерений многомерного шкалирования результатов морфологических промеров
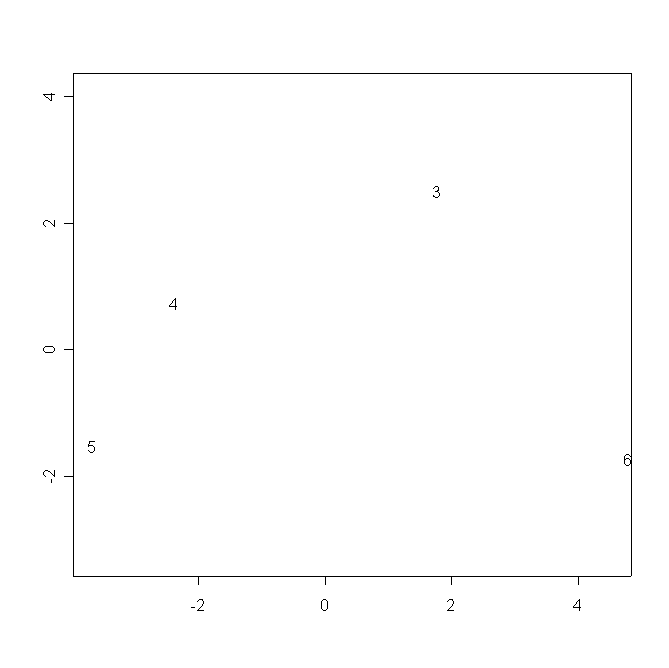
Условные обозначения:
См. условные обозначения к рис. 2б
В кластерном анализе морфологических характеристик можно наблюдать сходную картину (рис. 10).
Рисунок 10. Кластерный анализ морфологических характеристик популяций
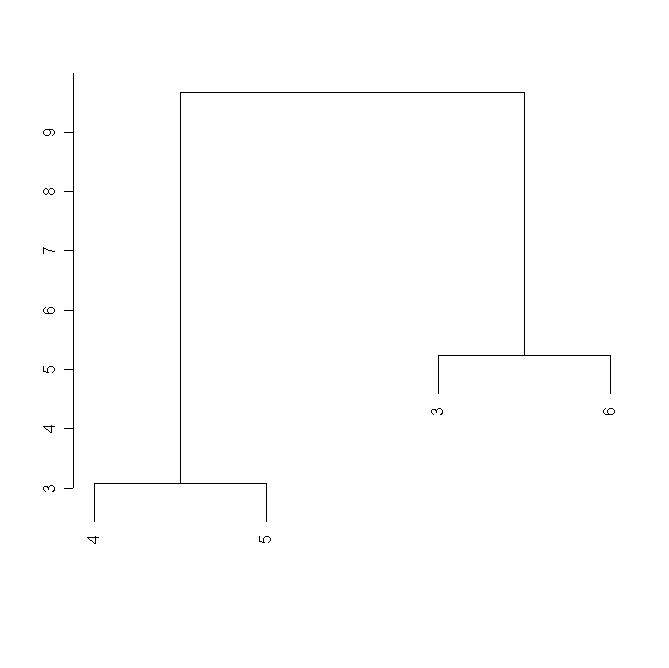
Условные обозначения:
См. условные обозначения к рис. 2б
Полученный методом суперимпозиции обобщенный поперечный срез срединной части черешка D. rotundifolia имеет сложную форму с ярко выраженной выпуклостью в средней части адаксиальной стороны (рис. 7а), а у D. anglica – округлотрехугольную форму (рис. 7в). У D. ×obovata поперечный срез срединной части черешка овальный, его адаксиальная сторона чуть более выпуклая, чем абаксиальная (рис. 7б). Поперечный срез срединной части черешка D. linearis имеет округлую форму (рис 1b).
Рисунок 7. Усредненная форма поперечного среза срединной части черешка исследованных видов Drosera
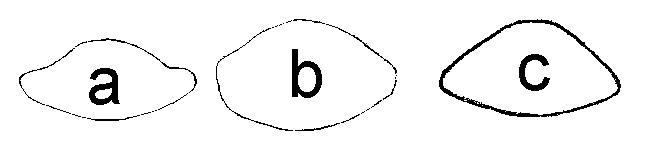
а – D. rotundifolia
б – D. ×obovata
в – D. anglica
При классификации отдельных особей методами геометрической морфометрии и классической морфометрии были получены сходные результаты. Растения разделялись согласно их видовой принадлежности, причем особи D. ×obovata занимали промежуточное положение между особями родительских форм, что полностью согласуется с литературными данными (Schnell, 1999). Форма поперечного среза срединной части черешка у D. anglica по данным геометрической морфометрии является промежуточной между формой такого среза у D. linearis и D. rotundifolia, что подтверждает данные C.E. Wood (1955) о гибридогенном происхождении D. anglica.
Надо отметить, что беломорская D. ×obovata ближе к D. rotundifolia, чем к D. anglica, как по форме поперечного среза черешка, так и по данным морфологических промеров. Это подтверждает полученные нами ранее на массовом морфологическом материале результаты (Волкова, 2002) и может свидетельствовать о преобладании возвратных скрещиваний в сторону D. rotundifolia и/или о значительном давлении отбора в сторону одной из двух возможных для наших росянок стратегий насекомоядности (Волкова и др., 2003).
Заслуживающим внимания фактом является объединение растений D. rotundifolia согласно району произрастания на основе их морфометрии, тогда как по признаку формы поперечного среза черешка такой обособленности не наблюдается. Растения D. rotundifolia из республики Карелия обладают более мелкими распростертыми листьями (длина и ширина листовой пластинки 5-7 мм, длина черешка 10-20 мм), в отличие от более крупных приподнятых над субстратом листьев D. rotundifolia из Тверской области (длина листовой пластинки 6-8 мм, ее ширина 8-10 мм, длина черешка 16-33 мм). Подобные различия, вероятно, вызваны разницей в климатических условиях. Форма поперечного среза черешка, таким образом, имеет меньшую географическую изменчивость, чем большинство метрических характеристик.
Характер распределения усредненных конфигураций и ЦО варьирует в зависимости от метода классификации. При использовании ЦО удовлетворительно интерпретируемую классификацию образцов можно получить лишь при многомерном шкалировании. Использование усредненных конфигураций как при многомерном шкалировании, так и в особенности при кластерном анализе данных, позволяет получить хорошо соответствующую литературным данным и результатам морфологических промеров картину. Необходимо учитывать, однако, что конфигурации для D. rotundifolia и D. linearis, заимствованные из работы C.E. Wood (1955), лишь формально могут считаться усредненными.
Изменчивость формы поперечного среза срединной части черешка у разных видов росянки наиболее сильно затрагивает не те части контура, которые должны были бы сильнее всего изменяться исходя из его видоспецифической формы. Этот факт позволяет согласиться с мнением И.Я. Павлинова (1999, 2000), о том, что методы геометрической морфометрии могут выявлять и трудно интерпретируемые (исходя из сложившихся представлений) закономерности.
Сбор большей части данных для настоящей работы проводился в рамках летних биологических практик Московской Гимназии на Юго-Западе (№1543). Мы благодарим руководителя практик С.М. Глаголева и всех участников практик, помогавших нам в полевой работе.
Волкова П.А. О морфологических отличиях трех видов росянки (Drosera L., Droseraceae) // Международная научная конференция по систематике высших растений, посвященная 70-летию со дня рождения чл.-корр. РАН, проф. В. Н. Тихомирова: Тез. докл. М., 2002. С. 29-30.
Волкова П.А., Кумскова Е.М., Шипунов А.Б. Зависимость морфофизиологических характеристик от успеха ловли насекомых и условий обитания у Drosera rotundifolia L., D. anglica Huds., D. obovata Mert. et Koch (Droseraceae) и Pinguicula vulgaris L. (Lentibulariaceae) // Бюллетень МОИП. Отд. биол. 2003. Т. 108. Вып. 1. С. 72-78.
Кирпичников М.Э. Сем. Droseraceae – Росянковые // Маевский П.Ф. Флора средней полосы европейской части СССР. 9-е изд. Л., 1964. С. 355.
Раменская М.Л., Андреева В.Н. Определитель высших растений Мурманской области и Карелии. Л., 1982. С. 251.
Павлинов И.Я., Волцит О.В., Россолимо О.Л. Анализ изменчивости формы методами геометрической морфометрии: демонстрация некоторых возможностей на примере гнатосомы клещей (Acari: Ixodes) и коренного зуба полевок (Mammalia: Alticola) // Журнал общей биологии. 1994. Т. 55. N 1. С. 110-118.
Павлинов И.Я. Анализ изменчивости формы третьего верхнего коренного у скальных полевок рода Alticola (Cricetidae) методами геометрической морфометрии // Зоологический журнал. 1999. Т. 78. N 1. С. 78-83.
Павлинов И.Я. Геометрическая морфометрия черепа мышевидных грызунов (Mammalia, Rodentia): связь формы черепа с пищевой специализацией // Журнал общей биологии. 2000. Т. 61. N 6. С. 583-600.
Павлинов И.Я., Микешина Н.Г. Принципы и методы геометрической морфометрии // Журнал общей биологии. 2002. Т. 63. N 6. С. 473-493.
Kores P.J., Molvray M., Darwin S.P. Morphomeric variation in three species of Cyrtostylis (Orchidaceae). // Systematic Botany. 1993. V. 18. N 2. P. 274-282.
Ray T.S. Landmark eigenshape analyis: homologous contours: leaf shape in Syngonium (Araceae) // Am. J. Bot. 1992. V. 79. N 1. P. 69-76.
Rohlf F.J. tpsUtil – TPS utility program. N.Y.: State Univ. at Stony Brook. 2000. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsRelw: relative warps. N.Y.: State Univ. at Stony Brook. Version 1.23. 2001a. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsDig. N.Y.: State Univ. at Stony Brook. Version 1.23. 2001b. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsSuper. N.Y.: State Univ. at Stony Brook. Version 1.12. 2003. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Schnell D. Drosera anglica Huds. vs Drosera х anglica: What Is the Difference? // Carnivorous Plants Newsletter. 1999. V. 28. N 4.
Wood C.E. Evidence for the hybrid origin of Drosera anglica // Rhodora. 1955. V. 57. P. 105-130.
Venables V.N., Smith D.N. and the R Development Core Team. An Introduction to R. London, 2002. 148 p.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |