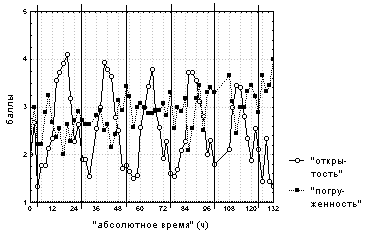
Вертикальными сплошными линиями отмечено 00:00 каждых суток наблюдений.
Большинству процессов, протекающих в живых организмах, свойственны ритмы эндогенной природы (Bunning, 1931). Многие авторы указывают на необходимость синхронизирующей стимуляции для возникновения ритмов в тканях растений и неспособность клеток зеленых растений синхронизировать друг друга (Уилкинс, 1964; Эмме, 1967; Уинфри, 1990). Существует и противоположное мнение о возможности передачи возбуждения от одной растительной клетки к другой различными способами (Штейн-Марголина, 1982). Важнейшая роль света для их синхронизации была неоднократно установлена разными авторами (Уилкинс, 1964; Эмме, 1967; Уинфри, 1990 и др.).
Поведение цветков кувшинки чисто-белой (Nymphaea candida Presl.) является одним из ярких примеров циркадных ритмов. Хорошо известно, что в ясную погоду цветки этого растения утром поднимаются на поверхность воды и открываются, а вечером погружаются под воду и закрываются (Кернер фон-Марилаун, 1895; Velde, 1986; Артамонов, 1989 и др.). Причины подобного поведения до сих пор окончательно не выяснены. Многие авторы указывают на то, что поведение цветка определяется освещенностью (Кернер фон-Марилаун, 1895; Эмме, 1967; Детари, Карциги, 1984; Уинфри, 1990). В. И. Артамонов (1989) считает, что поведение цветка в основном обусловлено тепературой воды и температурой воздуха, тогда как Б. Суини и Дж. Гастингс (1964) отрицают эту точку зрения. Предполагается, что изменение степени открытости и погруженности цветков вызваны изменением размера клеток вследствие изменения тургора (Кернер фон-Марилаун, 1895). Кроме того, причиной периодического изменения степени открытости цветка, по мнению А. Кернера фон-Марилауна (1895) и В.А. Штейн-Марголиной (1982), является неодинаковая интенсивность роста внутренней и внешней поверхности лепестков и чашелистиков во время развития цветка.
Существует множество гипотез о предназначении изменения степени открытости цветка в течение суток. По мнению разных авторов, это явление необходимо, чтобы удерживать в цветке насекомых-опылителей (Кернер фон-Марилаун, 1895) или "выбирать" опылителей определенных видов (Wiersema, 1988), предотвращать попадание росы на пыльцу (Кернер фон-Марилаун, 1895) или предотвращать теплопотери холодной ночью (Артамонов, 1989).
Ранее проведенные нами полевые наблюдения показали, что при естественном фоторежиме в средней полосе европейской части России изменение степени открытости и степени погруженности цветков зависит от природных факторов, имеющих суточный ход, таких как солнечная радиация и температура воды и воздуха (Волкова и др., 2002). В естественных условиях эти природные факторы тесно связаны между собой, поэтому не представлялось возможным установить, какой фактор является определяющим. В настоящей работе цветки наблюдались при различных фоторежимах в природных условиях с целью выявить основной природный фактор, обуславливающий поведение цветков.
Наблюдения проводили с 24 по 30 июня 2002 г. в течение 132 часов на оз. Молдино (Удомельский район Тверской области; 58 град. 15' с.ш., 34 град. 30' в.д.) и с 29 по 31 июля 2002 г. в течение 64 часов на оз. Тайное (Лоухский район республики Карелия; 66 град. 20' с.ш., 33 град. 20' в.д.). В Тверской области 15 цветков наблюдали при естественном фотопериоде (продолжительность светового дня 29 июня составляла 18 ч. 17 мин.). Также были проведены наблюдения за цветками в экспериментальных условиях. Пять цветущих растений в 00:00 28 июня в течение 3 секунд были освещены магниевой вспышкой. Еще 15 цветущих растений были почти полностью лишены солнечного света на все время наблюдений. Для этого вокруг растений была сооружена камера из светонепроницаемой полиэтиленовой пленки. Сторона камеры, обращенная к солнцу, и ее "крыша" были покрыты светоотражающей пленкой для предотвращения перегрева растений. Периодические измерения температуры воздуха показали, что существенного повышения температуры в камере по сравнению с открытым воздухом не наблюдалось. В темное время суток для наблюдения за цветками использовали фонарь с интерференционным "зеленым" светофильтром и малой интенсивностью света. Такое освещение практически не воспринимается растениями (Клешнин, 1954) и не нарушает условий эксперимента.
Наблюдения за 15 кувшинками в условиях отсутствия темновой фазы были проведены в Карелии (продолжительность светового дня 29 июля составляла 19 ч. 17 мин. при отсутствии темного времени суток).
К цветоносу каждого исследуемого растения была привязана плавающая на воде метка с индивидуальным номером. Подобная система мечения исключает путаницу цветков и сводит к минимуму влияние меток на жизнедеятельность растений. Для каждого цветка в течение всего срока наблюдений один раз в два часа отмечали степень его открытости и степень его погруженности и один раз в восемь часов -- стадию его развития. Эти характеристики оценивались визуально по пятибалльной шкале (табл. 1, 2).
| балл | открытость | погруженность |
| 1 | цветок полностью закрыт | цветок плавает на поверхности воды |
| 2 | цветок почти закрыт | цветок погружен на 1/3 своей высоты |
| 3 | цветок наполовину открыт | цветок погружен на 1/2 своей высоты |
| 4 | цветок почти открыт | цветок погружен на 2/3 своей высоты |
| 5 | цветок полностью открыт | цветок полностью погружен под воду |
| название стадии | состояние цветка |
| стадия бутона | цветок закрыт, чашелистики тесно прилегают друг к другу |
| пестичная стадия | цветок недавно раскрылся, пыльники незрелые, рыльцевый диск увлажнен |
| переходная стадия | пыльники созревают и склоняются над рыльцевым диском |
| тычиночная стадия | рыльцевый диск сухой, все пыльники созрели, активно продуцируется пыльца |
| конечная стадия | отцветание, пыльца не продуцируется, рыльцевый диск сухой |
В ходе каждого наблюдения измеряли атмосферное давление, температуру верхнего слоя воды и температуру воздуха. В Тверской области дополнительно визуально оценивали процент облачности, регистрировали относительную влажность воздуха при помощи аспирационного психрометра Ассмана и измеряли фотосинтетически активную солнечную радиацию, далее ФАР (интервал длин волн 400-710 нм; Воскресенская, 1965). Измерения ФАР проводили при помощи цветного пиранометра ГГО М-80 в комплекте с гальванометром ГСА-1 (Махоткина, 1983). Для каждого фоторежима вычислили "средний балл степени открытости" цветков (среднее арифметическое баллов открытости всех цветков за одно наблюдение) и "средний балл степени погруженности" цветков (среднее арифметическое баллов погруженности всех цветков за одно наблюдение) для получения представления об общих тенденциях поведения цветков при данном фоторежиме.
Сведения о длине светового дня были получены при помощи компьютерной программы астрономических расчетов XEphem (Downey, 2000).
Мы анализировали поведение цветков, находящихся на пестичной, переходной и тычиночной стадиях, поскольку именно на этих стадиях цветок выполняет свою репродуктивную функцию (Velde, 1986; Wiersema, 1988). Провели визуальный анализ графиков зависимости степени открытости цветков и степени их погруженности от "абсолютного времени", то есть времени, прошедшего с момента начала наблюдений (рис. 1-4). Рассчитали коэффициенты корреляции между степенью открытости цветков, степенью их погруженности, атмосферным давлением, температурой верхнего слоя воды, температурой воздуха, процентом облачности, относительной влажностью воздуха и ФАР. Распределение данных было отличным от нормального, поэтому мы использовали непараметрические коэффициенты ранговой корреляции Спирмана (Зайцев, 1984).
Данные обрабатывали при помощи пакета STATISTICA for Windows (StatSoft, Inc., 1999).
"Средняя степень открытости" и "средняя степень погруженности" цветков с разной силой коррелируют со степенью открытости и степенью погруженности соответственно каждого из цветков (R=0,3-0,7). Таким образом, общие тенденции поведения всех цветков адекватно описываются этими усреденнными показателями.
Наблюдалась четкая ритмика изменения "средней степени открытости" цветков с 24-х часовым периодом на фоне постепенного увеличения "средней степени погруженности" со слабо выраженной суточной ритмикой, что связано, вероятно, с отцветанием растений. Цветки каждые сутки были открытыми с 10:00-12:00 до 14:00-16:00 и были закрытыми с 18:00-0:00 до 8:00, при этом все открытые цветки плавали на поверхности воды (рис. 1).
Вертикальными сплошными линиями отмечено 00:00
каждых суток наблюдений.
При анализе поведения отдельных цветков заметен больший разброс времени нахождения цветков в открытом состоянии (с 8:00-12:00 до 14:00-22:00). Кроме того, у каждого из исследуемых цветков на разных стадиях развития наблюдались различные отклонения от типичного поведения, не зависящие от времени суток или региструемых погодных условий. Около 75% цветков неоднократно полностью или наполовину погружалось под воду в открытом или полуоткрытом состоянии, для половины цветков были отмечены частые изменения степени погруженности или всплытие на поверхность воды в темное время суток.
Из описания поведения цветков видно, что при естественном для средней полосы фоторежиме наблюдалась слабая обратная зависимость "средней степени открытости" цветков от их "средней степени погруженности". Такая зависимость (R=-0,3... 0,5) была отмечена лишь для 30% цветков, содержащихся при данном фоторежиме.
"Средняя степень открытости" цветков была прямо пропорциональна (R=0,6) интенсивности ФАР, тогда как для их "средней степени погруженности" наблюдалась обратная зависимость (R=-0,5). Подобная зависимость, хотя и выраженная менее ярко, наблюдалась для степени открытости (R=0,3-0,5) 60% цветков и для степени погруженности (R=-0,4... -0,5) 20% цветков, наблюдаемых при данном фоторежиме. "Средняя степень открытости" цветков также была прямо пропорциональна (R=0,4-0,5) температуре верхнего слоя воды и температуре воздуха. Подобная зависимость отмечена для 60% цветков, наблюдаемых при данном фоторежиме.
Визуальный анализ зависимости степени открытости цветков и степени их погруженности от "абсолютного времени" (рис. 2) позволил отметить следующие особенности в поведении этих цветков.
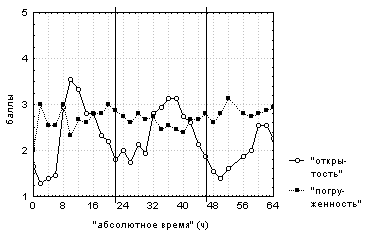
Вертикальными сплошными линиями отмечено 00:00
каждых суток наблюдений.
Более половины исследованных цветков имеют отличное от описанного в литературе поведение. В частности, наблюдали кратковременное (не более чем на один час) открывание плавающих на поверхности цветков, кратковременное погружение под воду полуоткрытых и полностью открытых цветков и длительное (около 12 часов) нахождение в полупогруженном состоянии открытых цветков. Вышеописанное поведение было отмечено для цветков на разных стадиях развития в различное время суток, что позволяет говорить об эндогенной природе таких отклонений от описанного в литературе поведения цветков. Лишь менее половины исследованных цветков открывалось ежедневно на срок от двух до 12 часов, при этом открытый цветок практически во всех случаях не был погружен под воду. Остальные цветки открывались в один или в два дня из трех дней наблюдений за ними, также не будучи погруженными под воду. Все цветки открывались в разное время суток и на разные сроки, что не зависело ни от стадии развития цветка, ни от регистрируемых погодных условий.
Интересно, что подавляющее большинство исследованных цветков мало времени проводило полностью погруженными под воду и б/'ольшую часть времени находилось на поверхности воды. При этом цветки, как правило, были открытыми лишь около одной четверти общего времени наблюдений.
По данным корреляционного анализа лишь у 40% исследованных цветков наблюдалась обратная зависимость (R=-0,5... -0,8) степени открытости цветка от степени его погруженности. "Средняя степень открытости" цветков коррелирует (R=0,4-0,9) со степенью открытости каждого из цветков, наблюдаемых при данном фоторежиме. "Средняя степень погруженности" цветков коррелирует (R=0,4-0,6) со степенью погруженности лишь половины цветков. Таким образом, "средняя степень открытости" адекватно характеризует изменение степени открытости каждого из наблюдаемых при данном фоторежиме цветков, тогда как "средняя степень погруженности" описывает лишь общие тенденции изменения степени погруженности каждого из цветков.
Визуальный анализ графиков зависимости "средней степени открытости" цветков и их "средней степени погруженности" от времени, прошедшего с момента начала наблюдений, позволил выявить общие тенденции поведения цветков. Как правило, цветки находятся на поверхности воды. При этом им свойственно ежедневно раскрываться с 8:00 до 12:00-14:00. "Средняя степень открытости" цветков изменяется с 24-х часовым периодом, тогда как изменения "средней степени погруженности" не имеют четкой периодичности. Однако, как видно из описания поведения каждого из наблюдаемых цветков, из общего правила существует множество исключений и, кроме того, поведение разных цветков слабо скоррелировано между собой.
Степень открытости всех цветков при данном фоторежиме положительно связана (R=0,5-0,7) с температурой верхнего слоя воды и с температурой воздуха.
"Средняя степень открытости" и "средняя степень погруженности" цветков коррелируют соответственно со степенью открытости (R=0,5-0,8) и степенью погруженности (R=0,5-0,6) лишь половины всех цветков, содержащихся в темноте. Таким образом, усредненные показатели описывают лишь общие тенденции поведения цветков при этом фоторежиме.
В течение всего времени наблюдений цветки то плавали на поверхности воды, то наполовину погружались под воду вне зависимости от времени суток и "абсолютного времени" (рис. 3).
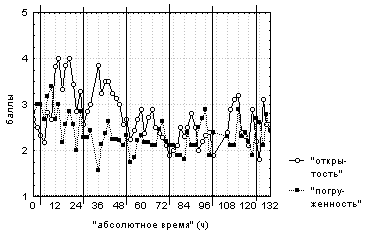
Вертикальными сплошными линиями отмечено 00:00
каждых суток наблюдений.
Первые двое суток наблюдений цветки открывались днем с 8:00 до 14:00-18:00, а ночью оставались наполовину открытыми. В последующие трое суток наблюдений цветки постоянно были полуоткрытыми (рис. 3), что связано, вероятно, с отцветанием большинства исследуемых растений и характерно для 70% исследуемых цветков. Изменения "средней степени открытости" цветков имеют период, близкий к 24-м часам, тогда как изменения "средней степени погруженности" цветков не имеют четкой периодичности. Анализ поведения отдельных цветков демонстрирует существенно меньшую синхронность изменения степени открытости (цветки открыты с 4:00-12:00 до 14:00-24:00). Угроза попадания воды внутрь цветка вследствие погружения под воду незакрытого цветка неоднократно возникала у 70% исследуемых цветков.
Это описание демонстрирует отсутствие типичной для естественного фоторежима обратной зависимости степени открытости цветков от их степени погруженности, что подтверждается и корреляционным анализом данных.
Четкой зависимости поведения цветков от региструемых погодных условий обнаружено не было.
"Средняя степень открытости" и "средняя степень погруженности" цветков коррелируют соответственно со степенью открытости (R=0,5-0,9) и степенью погруженности (R=0,6-0,8) каждого из цветков, подвергнутых вспышке света в темное время суток. Таким образом, поведение всех цветков при этом фоторежиме адекватно описывается усреденнными показателями.
Мы не имеем достаточно данных для того, чтобы с уверенностью судить о периодичности изменения "средней степени открытости" и "средней степени погруженности" цветков. Однако имеющиеся данные не противоречат предположению о наличии 24-х часового периода изменения "средней степени открытости" цветков и при этом фоторежиме.
Цветки были открытыми в следующий после освещения день с 10:00 до 16:00, при этом они были полностью погружены в воду. Ночью (с 0:00 до 4:00-8:00) первых двух суток после освещения цветки были полностью закрыты и погружены под воду. В остальное время цветки были наполовину открытыми и наполовину или полностью погруженными под воду вне зависимости от стадии развития цветка, времени суток и регистрируемых погодных условий (рис. 4). Для отдельных цветков также было характерно нахождение под водой в открытом состоянии или всплытие на поверхность воды закрытого цветка.
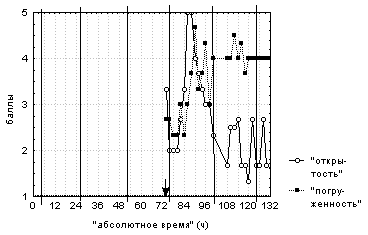
Вертикальными сплошными линиями обозначено 00:00
каждых суток наблюдений. Стрелка указывает на
момент вспышки света.
Это описание демонстрирует отсутствие типичной для естественного фоторежима обратной зависимости степени открытости цветков от их степени погруженности, что подтверждается и корреляционным анализом данных. Поведение цветков во второй половине срока наблюдений за ними, вероятно, обусловлено их отцветанием.
Интересно отметить, что цветки б/'ольшую часть времени были полузакрытыми и находились на поверхности воды и/или в открытом состоянии лишь очень небольшое время.
Корреляционный анализ данных свидетельствует о том, что "средняя степень открытости" этих цветков прямо зависит от интенсивности ФАР (R=0,8) и от температуры верхнего слоя воды (R=0,5).
Общие тенденции изменения степени открытости цветков, наблюдаемые в средней полосе при естественном фоторежиме, проявляются и при других фоторежимах. Изменение степени открытости цветков при всех фоторежимах имеет 24-х часовой период. Это свидетельствует о том, что изменение степени открытости цветков является свободно протекающим эндогенным ритмическим процессом (Эмме, 1967; Детари, Карциги, 1984).
Степень погруженности изменяется с плохо заметной периодичностью, которая, как правило, выражена значительно слабее нелинейного тренда. Лишь при естественном фоторежиме в средней полосе наблюдалась описанная в литературе обратная зависимость степени открытости цветков от степени их погруженности. Отсюда следует, что изменению степени погруженности цветков свойственна слабовыраженная ритмичность экзогенной природы (Эмме, 1967).
У многих представителей семейства Nymphaeaceae наблюдается изменение степени открытости у постоянно плавающего на поверхности воды цветка (Sculthorpe, 1967). Степень открытости цветка определяет его способность к опылению, то есть к выполнению основной функции цветка. Вероятно, это обстоятельство служит причиной наличия эндогенных ритмов изменения степени открытости, в отличие от степени погруженности. Очевидно, что изменение степени открытости цветка обеспечивается активностью клеток околоцветника, тогда как степень погруженности цветка обусловлена состоянием клеток его цветоноса. Отсутствие взаимной синхронизации у растительных клеток, а также важнейшая роль света для их синхронизации были неоднократно установлены разными авторами (Уилкинс, 1964; Эмме, 1967; Уинфри, 1990 и др.). Вероятно, степень открытости цветка и степень его погруженности изменяются независимо друг от друга и синхронизируются изменением интенсивности освещенности. Этот вывод не согласуется с мнением В.А. Штейн-Марголиной (1982) в отношении возможности синхронизации клетками околоцветника и цветоноса друг друга.
В случае несоответствия фотопериода естественному для средней полосы привычная нам связь между степенью открытости цветка и степенью его погруженности исчезает. Наблюдаемая нами как в этом году, так и в предыдущие годы (Волкова и др., 2002) положительная связь степени открытости цветков с температурой верхнего слоя воды и температурой воздуха, вероятно, вызвана суточным ходом этих факторов. Это подтверждается отсутствием такой связи в эксперименте без световой фазы. Зависимость поведения цветков от относительной влажности воздуха не обнаружена, что согласуется с полученными нами ранее результатами (Волкова и др., 2002) и противоречит мнению А. Кернера фон-Марилауна (1895). Также не было выявлено влияния атмосферного давления на ритмику изменения степени открытости и степени погруженности цветков.
Нам представляется, что единственной целью периодического закрывания цветка может являться повышение эффективности опыления цветка насекомыми согласно А. Кернеру фон-Марилауну (1895) и Wiersema (1988).
Настоящая работа проводилась на Молдинской Биостанции Московской Гимназии на Юго-Западе, а также в ходе Беломорской экспедиции Гимназии, проходившей под руководством к.б.н. С.М. Глаголева. Ценные указания при подготовке рукописи сделаны к.б.н. А.Б. Шипуновым, к.б.н. Д.Д. Соколовым и проф. И.А. Шульгиным. Большую помощь в статистической обработке данных оказал проф. В.Н. Тутубалин. В проведении полевых наблюдений, организованных А.Б. Шипуновым, активнейшее участие принимали преподаватель Гимназии А.Н. Квашенко, аспирантка биологического факультета МГУ Я.В. Косенко и ученицы Гимназии О. Бочкова, О. Васильева, Т. Волкова, Е. Кольчугина и Е. Пескова. Метеорологическое оборудование и консультации по его использованию были предоставлены старшим научным сотрудником Метеообсерватории МГУ к.г.н. О.А. Шиловцевой.
Артамонов В.И. Зеленые оракулы. М., 1989.
Волкова П.А., Сонина С.И., Шипунов А.Б. Особенности поведения цветков кувшинки чисто-белой (Nymphaea candida Presl.) на оз. Молдино (Тверская область) // Бюллетень МОИП. Отд. биологич., 2002. Т. 107, Вып. 5. С. 57-63.
Воскресенская Н.П. Фотосинтез и спектральный состав света. М., 1965. С. 6, 161, 210-215, 256.
Детари Л., Карциги В. Биоритмы. М., 1984. С. 19, 25-26.
Зайцев Г.Н. Математическая статистика в экспериментальной ботанике. М., 1984. С. 130-138.
Кернер фон-Марилаун А. Жизнь растений. Т. 2. Спб., 1895. С. 202-205.
Клешнин А.Ф. Растение и свет. Теория и практика светокультуры растений. М., 1954. С. 11-12, 126-134.
Махоткина Е.Л. Цветные пиранометры. 1983. Вып. 456. С. 71-77.
Суини Б., Гастингс Дж. Влияние температуры на суточные ритмы // Биологические часы. М., 1964. С. 152-155.
Штейн-Марголина В.А. Некоторые аспекты движения листьев // Успехи современной биологии, 1982. Т. 94. вып. 1(4). С. 111-125.
Уилкинс М. Влияние света на ритмы растений // Биологические часы. М., 1964.
Уинфри А.Т. Время по биологическим часам. М., 1990.
Эмме А.М. Биологические часы. Новосибирск. 1967. С. 12, 16, 72-93.
Bunning E. Untersuchungen uber die automen tagesperiodischen Bewedungen der Primarblatter von Phaseolus multiflorus // Jahrb. Wiss. Bot. 1931. Vol. 75. P. 439-480.
Downey E.C. Xephem. Astronomy program. Version 3.2.2. 2000. [Electronic resource]. Mode of access: http://www.clearskyinstitute.com/xephem/.
Sculthorpe C.D. The biology of aquatic vascular plants. London. 1967.
StatSoft, Inc. STATISTICA for Windows [Computer program manual]. Tulsa, OK. 1999.
Velde, van der G. Developmental stages in the floral biology s. l. of dutch Nymphaeaceae (Nymphaea alba L., Nymphaea candida Presl., Numphar lutea (L.) Sm.) // Acta Bot. Neerl. 1986. Vol. 35. N 2. P. 111-113.
Wiersema H.J. Reproductive biology of Nymphaea (Nymphaeaceae) // Annals of the Missouri Botanical Garden. 1988. Vol. 75, N 3. P. 796-804.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |