Revealing the total species richness for the given area is well-known to be impossible, although this is a basic attribute for many models of community structure and biodiversity investigations (Gotelli, Colwell, 2001). Sparse investigations of the total species number revealing were based mainly on zoological material (Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005 and references therein, but see Miller, Wiegert, 1989; Scherbakov, 2003). This situation could be caused by greater urgency of revealing total species number for zoologists then for botanists, because direct observation of animals is more complex. However, revealing of the perfect flora also appears to be a challenge mainly because of the existence of some species in undetectable condition (e.g. as seed bank or underground organs) in certain seasons or even years and also because of accidental overlook of the detectable species (Scherbakov, 2003). Thus, this problem deserves more detailed investigation. It is more convenient to study the process of perfect flora revealing on the objects with clear natural borders, such as islands and lakes.
Equilibrium model of island biogeography (MacArthur, Wilson, 1967) has been tested on many objects since its publication. Sparse investigations were focused on the dynamics of islands flora. These investigations were based on the comparison of floristic lists, compiled for the same island in different years. Differences between these lists were treated as changes in island flora (Abbot, Black, 1980).
Thus, investigators of island flora dynamics actually equated the floristic lists compiled during one field season with perfect island flora what is evidently incorrect (Scherbakov, 2003). Some researchers did not rise this question at all (Reimanek, Rejmankova, 2002), others did not realize the problem, supposing that "careful searching" can reveal the perfect island flora (Abbot, Black, 1980), the thirds recognized the possibility of separate species overlooking (Vidal et al., 1998; Shipunov, Abramova, 2006), although the authors of the second paper supposed that "this factor did not play any significant role in our case".
The question about exact species that had been usually overlooked is also interesting. B. Herloff (2003) showed that solitary individuals growing in hard-hitting places had been usually overlooked. A. Shipunov and L. Abramova (2006) suggested that it was possible to compile the list of "the frequently overlooked species", which "had almost nothing common" with the lists of appearing or disappearing species on the islands. All this information let one suppose that the problem of species overlooking during floristic investigations in the islands is mainly imaginary. However, changes of islands flora were shown to happen owing to the species with low abundance (Shipunov, Abramova, 2006), that could be overlooked with great probability (Herloff, 2003). It remains unclear for us, for example, what part of “appeared” on the island species has been simply overlooked during the previous investigation.
These considerations lead us to the necessity of the investigation of the detectable plant species revealing on the two type of territories with clear natural borders (i.e. islands and lakes) with two methods that correspond the specificity of chosen objects (i.e. routes and tentative sites methods respectively).
We have investigated four islands of the Kandalaksha gulf of the White sea. Two islands of the Kuzakotskaya bay (Pokormyozhnyj and Kokoikha) were investigated in the beginning of July 2003, two more islands of the Chupa bay were studied in the end of July 2006. Each island was investigated by two independent research groups. We calculated four coefficients that were used by A. Shipunov and L. Abramova (2006) to characterize the changes of islands flora for each pair of floristic lists: the coefficient of disappearing (the fraction of disappeared species from the total species number of the island), the coefficient of appearing (the fraction of appeared species), the coefficient of stability (the fraction of constant species; describes the changes in species composition) and the coefficient of tendency (the difference between the two first coefficients, describes the changes of species richness). Simultaneous floristic descriptions were treated as descriptions made in the different years. For example, the species was treated as “disappeared” if it was overlooked by one of the groups. We also compared our floristic lists for the two islands of the Chupa bay with analogous lists, got in the beginning of August 2001 (White sea expeditions…, 2001; Abramova et al., 2003) to reveal the changes in islands flora during these five years. The investigations of islands flora were made by the method of transects with rounding of the seashore part. The research group went through the island frontally along its maximum length, then on the seashore, then along its maximum width and again on the seashore to the debarkation point. The most interesting places on the island (e.g. bogs) were studied more detailed (Shipunov, Abramova, 2006).
We calculated Cohen’s kappa to estimate the degree of agreement between pairs of simultaneous floristic descriptions made by different research groups (Petrie, Sabin, 2000) in R statistical environment (R Development Core Team, 2005).
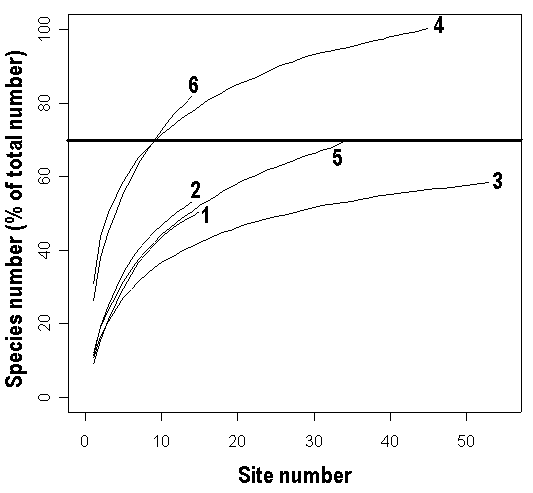
Flora of shores of six lakes (three in the Northern Karelia and three in the Tver region) was investigated to study the process of perfect flora revealing. We laid several sample sites on the shore of each lake and studied flora for each of them. Each site was 20 m lengthwise and had width from the beginning of the aquatic plants zone to 1 m away from the water’s edge. We used different algorithms of sample sites arrangement. Coastal line of one of the lakes (lk. Blizhnee) was parceled out, on the other lake (lk. Moldino) two parts of coastal line on the opposite shores were divided on the sites, on the other three lakes small number (14-34) of sample sites were uniformly distributed along all the costal line. Then “accumulation curves” (i.e. the curves of dependence of the total revealed species number on the site number) were plotted for each lake (Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005). We used averaged accumulation curves (Fig. 1), i.e. the result of 100 curves averaging, plotted after random permutations of sites number (rarefaction curves: Gotelli, Colwell, 2001). Total species number was estimated as asymptotic value for the Clench function fitted to the rarefaction curve. We have chosen the Clench function, because it was the best for fitting our rarefaction curves, i.e. the sum of squared residuals was minimal among all functions, optimal for this aim (Hortal, 2006). We also estimated total species number in the alternative way for the reference, via non-parametric estimator (Chao2: Hortal et al., 2006), calculated in the EstimateS 7.5.1 software (Colwell, 2005). All other calculations and graphs creation were made in the R statistical environment (R Development Core Team, 2005), used R-script for rarefaction curves building is available on http://herba.msu.ru/shipunov/software/r/r-en.htm.
Cohen’s kappa was 0.7 for the Kuzokotskaya bay islands and 0.9 for the Chupa bay islands, what was the evidence of good and excellent agreement between simultaneous floristic studies accordingly (Petrie, Sabin, 2000).
The part of overlooked by one of the research groups species (the analog of the coefficients of disappearance and appearance) was 5-15% from the total species number for the island. "The coefficient of tendency", calculated for simultaneous floristic studies of the same island was 5-7% and "coefficient of stability" was 74-90% (Table 1).
Table 1. Values of the used coefficients for simultaneous island investigations
| Island name | Total species number | "Coefficient" of... | |||
| disappearance | appearance | tendency | stability | ||
| Pokormyozhnyj | 180 | 0.11 | 0.06 | 0.00 | 0.90 |
| Kokoikha | 131 | 0.05 | 0.05 | 0.05 | 0.83 |
| Utichij | 97 | 0.15 | 0.09 | 0.07 | 0.74 |
| Chernyshov | 90 | 0.11 | 0.09 | 0.02 | 0.80 |
The species that had appeared or disappeared on the Kem-ludy islands (situated approximately between Kuzokotskaya and Chupa bays) are almost absolutely different from species overlooked by one of the groups on the Kuzokotskaya bay islands (two common species Carex buxbaumii and Scutellaria galericulata -- 4% of “Kem-Ludy list”) as it was correctly pointed out by A. Shipunov and L. Abramova (2006) and with species overlooked in the Chupa bay islands (only Melampyrum sylvaticum).
The species that had appeared or disappeared on the Chupa bay islands are also almost different from overlooked ones by one of the groups on the Kuzokotskaya bay islands (two common species: Achillea millefolium and Carex globularis). But at the same time more then a half of appeared on the Chupa bay islands species (52%) were overlooked by one of the research groups (6 species per each group).
Overlooked species usually were not very rare on the island (more frequent then “solitary”), had different life forms (trees -- Alnus incana, shrubs -- Salix caprea, Rosa majalis and others, numerous herbaceous plants), grew in the different habitats (from the Sphagnum bogs in the central part of the island -- Rubus chamaemorus -- to the middle littoral -- Zostera marina) and had different appearance (from the plain Poa pratensis to the bright Coronaria flos-cuculi).
Different species were usually overlooked on the close islands with similar flora. There were few exceptions: Botrychium boreale, Corallozhiza trifida, Goodyera repens, Juncus athrofuscus, Lycopodium annotinum, Carex globularis, C. aquatilis and Galeopsis bifida that were overlooked by different research groups on the several islands of the Kuzokotskaya and Chupa bays (Table 2).
Table 2. Overlooked, disappeared and appeared species (in alphabetical order)
| Overlooked species (2006) | Chupa bay islands: 2001 – 2006 | ||
| Kuzokotskaya bay islands | Chupa bay islands | disappeared species | appeared species |
| Achillea millefolium, Agrostis gigantea, Alnus incana, Angelica sylvestris, Botrychium boreale, B. lunaria, Calamagrostis purpurea, Carex buxbaumii, C. globularis, C. lasiocarpa, C. rotundata, Corallorhiza trifida, Dactylorhiza maculata, Deschampsia caespitosa, Equisetum pratense, Galeopsis bifida, Glaux maritima, Goodyera repens, Hippuris tetraphylla, Juncus artrofuscus, Lathyrus palustris, Lycopodium annotinum, Moneses uniflora, Oxycoccus microcarpus, Poa nemoralis, P. glauca, Ranunculus reptabundus, Rosa majalis, Rumex aquaticus, Salix glauca, S. hastata, S. lapponum, Scutellaria galericulata, Selaginella selaginoides, Stellaria crassifolia | Agrostis stolonifera, Antennaria dioica, Calamagrostis neglecta, Carex aquatilis, C. brunescens, C. glareosa, C. globularis, C. mackenziei, C. rariflora, C. rostrata, C. salina, Cerastium holosteoides, Cochlearia groenlandica, Conioselinum tataricum, Coronaria flos-cuculi, Erysimum hieracifolium, Equisetum fluviatile, E. sylvaticum, Galeopsis bifida, Gymnocarpium dryopteris, Heracleum sibiricum, Hieracium spp., Huperzia selago, Lycopodium annotinum, L. complanatum, Melampyrum sylvaticum, Parnassia palustris, Polygonum viviparum, Poa pratensis, Polypodium vulgare, Ranunculus polyanthemos, Rubus chamaemorus, Salix caprea, Sedum acre, Stellaria media, Zostera marina | Achillea millefolium, Agrostis tenuis, Comarum palustre, Eleocharis uniglumis, Epilobium palustre, Polygonum boreale, Potentilla egedii, Stelaria graminea, Vicia sepium | Agrostis gigantea, A. stolonifera, A. straminea, Antennaria dioica, Aster tripolium, Campanula rotundifolia, Carex aquatilis, C. glareosa, C. globularis, C. mackenzii, C. salina, Heracleum sibiricum, Huperzia selago, Parnassia palustris, Poa pratensis, Pyrola chlorantha, P. minor, Stellaria media, Vicia cracca, Festuca rubra, Ruppia maritima, Sagina nodosa, Sedum acre, Solidago virgaurea, Triglochin palustre, Zostera marina |
The number of sample sites that were needed to reveal lake flora on the preliminary level (all species with abundance “occasionally” and more, 70% of the perfect flora: Scherbakov, 2003), almost did not depend on the lake size and on the region (Fig. 1, Table 3). This was supported by the significant decreasing of the coastal line part that should be investigated with this aim while lake size increased (Table 3). Thus, one should investigated 9-25 sample sites (200-500 m of coastal line) on the average to reveal the lake flora on the preliminary level not depending on the lake size.
Table 3. Main features of the investigated lakes
| Lake name | Length of coastal line (m) | Investigated * | Total species number ** | Revealed 70% of total species number* | Chao2 **** |
| Tver region | |||||
| Olshevo | 6 350 | 15 (5) | 153 | 19 (6)*** | (102) 109±5 (124) |
| Chepolshevo | 5 000 | 14 (6) | 199 | 14 (6) | (156) 176±14 (214) |
| Moldino | 26 200 | 53 (4) | 169 | 18 (1) | (163) 179±12 (212) |
| Karelia | |||||
| Blizhnee | 900 | 45 (100) | 45 | 11 (24) | (43) 45 ±3 (57) |
| Dolgoe | 4 320 | 34 (16) | 166 | 21 (10) | (162) 192±26 (270) |
| Nizhnee Pulongskoe | 9 540 | 14 (3) | 124 | 9 (2) | (44) 62±19 (129) |
* -- number of sites (length of coastal line, % of total length)
** -- calculated as limit of Clench model (Soberon, Llorente, 1993)
*** -- calculated by extrapolation of Clench model (Soberon, Llorente, 1993)
**** -- nonparametric species richness estimator (Colwell, 2005; Hortal et al, 2006): (95% confidence interval low bond) estimator value ±SD (95% confidence interval upper bond)
Figure 1. Mean accumulation curves for different lakes
1 – lk. Olshevo, 2 – lk. Chepolshevo, 3 –lk. Moldino, 4 – lk. Blizhnee, 5 – lk. Dolgoe, 6 – lk. Nizhnee Pulongskoe. Total species number for the lake, taken as 100%, calculated as limit of Clench model (Soberon, Llorente, 1993).
Our investigations of perfect lake flora revealing showed that inventory of detectable species of relatively poor partial floras of northern taiga even on the preliminary level demands significant efforts. This result was supported also by large differences (up to 15% of total species number) between simultaneous floristic studies of the same island.
Simultaneous independent floristic investigations of islands showed that species were overlooked by pure accident, without any dependence on their appearance or habitat. That is why we consider analysis of single species dynamics on the island to be fruitless as it can be caused by species overlooking during the earlier investigation (one would observe species “appearance”) or during the later investigation (species “disappearance”). From the other hand, complex analysis of species dynamics (after data aggregating for the several islands), as it was done by A. Shipunov and L. Abramova (2006), is, in our opinion, a successful approach. According to our data one is likely to overlook different species on the different (although quite similar) islands. Thus, the consequences of accidental species overlooking “self-annihilate” after the data aggregation for a number of similar islands
Thus, floristic description can be treated as a sample from the universe, which is represented by perfect flora of the given area. That is why the comparison of check-lists (in particular during revealing of island flora dynamics) is to be conducted using the logics of statistical samples comparison. Statistical significance of the difference between samples is determined by comparison of the probability of this difference existence with fixed critical value. In the investigations of island flora dynamics probability of significant changes in island flora can be expressed by the coefficients, which are to characterize these changes (Shipunov, Abramova, 2006). Coefficient values for two independent simultaneous floristic investigations of the same island (equivalent of two statistical samples from one universe, differing only by chance) should be treated as critical values. This pair of investigations will differ only due to accidental species overlooking by one of the research groups. Our investigations showed that critical value for the coefficients of appearance and disappearance can be established as 0.15, for the coefficient of stability – 0.7 and for the module of tendency coefficient – 0.1. This means that changes of island flora reflect real flora dynamics and not the investigation error if value of stability coefficient does not exceed its critical value and values of the other coefficients are greater then their critical values.
Usage of the suggested critical values lets reveal real (i.e. not caused by accidental species overlooking) changes in island flora. One should note that these changes could be caused by the differences in weather conditions (some plants can be in undetectable condition in the unfavorable years) and not by flora dynamics in the context of equilibrium model of island biogeography (MacArthur, Wilson, 1967). Causes of the noticed flora dynamics should be probably revealed in series (not less then four) successive investigations of island flora. However, besides great laboriousness of this method, the possibility of influence of researches activity on the island flora is substantial disadvantage of the method.
Noticed regularities of detectable species revealing definitely can not be automatically spread to all floristic investigations. We expect that concrete parameters of these regularities (for example, critical values of coefficients used to characterize island flora dynamics) will depend on many factors such as size of the investigated area, native zone, researchers’ qualification etc. For example, agreement between simultaneous investigations on the Chupa bay islands was higher than on the Kuzokotskaya bay islands. From the one hand, this can be explained by smaller size of the Chupa bay islands. From the other hand, investigations of these islands were made by the members of the White sea expedition of Moscow South-West high school with extensive experience of collaborative floristic work, while the islands of Kuzokotskaya bay were investigated by poorly coordinated research groups.
We do not pretend to universality of the described patterns of the process of flora revealing. One should underline only that such investigations are necessary for the determination of sensitivity of the selected method of floristic investigations. Solving of concrete floristic problems (in particular, connected with comparison of floras of different areas or in different periods) without any information about “sensitivity” of the used method is similar to the measurements with ruler with unknown scale factor. Such data have small scientific value. Estimation of the fraction of revealed species and total species number prediction on the given area with help of the accumulation curves is essential for the inventory works, what was repeatedly pointed out by a number of researchers (Miller, Wiegert, 1989; Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005). The similar problem of the estimation of complex method sensitivity became relevant recently in the completely different investigation area – non-direct DNA analysis. In these methods human factor plays an important role as it does in the floristic researches. Molecular biologists insist on determination of the method sensitivity in every work and publishing it to solve this problem (Bonin et al., 2004). We think that florists should follow this example.
Abbott J., Black R. Changes in species composition in floras on islets near Perth, Western Australia // Journ. Biogeogr. 1980. Vol. 7. N. 4. P. 399–410.
Abramova L.A., Rimskaya-Korsakova N.N., Shipunov A.B. Comparative investigations of flora of the Kiv bay, Chupa bay and Keretskij archipelago (Kandalaksha gulf, White sea) // N.A. Pertsov conference. Vol. 9. Moscow, 2003. P. 22-33 [In Russian].
Bonin A., Bellemain E., Eidesen P.B., Pompanon F., Brochmann C., Taberlet P. How to track and assess genotyping errors in population genetics studies // Molecular Ecology 2004. Vol. 13. P. 3261-3273.
Colwell R.K. EstimateS: Statistical estimation of species richness and shared species from samples. 2005. Version 7.5.1. [Electronic resource]. Mode of access: http://purl.oclc.org/estimates.
Diaz-Frances E., Soberon J. Statistical estimation and model selection of species accumulation curves // Conservation Biology. 2005. Vol. 19, N 2. P. 569-573.
Gotelli N.J., Colwell R.C. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness // Ecology Letters. 2001. Vol. 4. P. 379–391.
Herloff B. Floristic changes during the twentieth century in the southern archipelago of Goeteborg, SW Sweden // Svensk Bot. Tiddskr. 2003. Vol. 97. P. 3–14.
Hortal J., Borges P.A.V., Gaspar C. Evaluating the performance of species richness estimators: sensitivity to sample grain size // Journal of Animal Ecology. 2006. Vol. 75. P. 274-287.
McArthur R.H., Wilson O.E. The theory of island biology. Princeton, New Jersey, 1967. 203 p.
Miller R.I., Wiegert R.G. Documenting completeness, species-area relations and the species-abundance distribution of a regional flora // Ecology. 1989. Vol. 70, N 1. P. 16-22.
Petrie A., Sabin C. Medical statistics at a glance. Blackwell science, London. 2000. 138 p.
R Development Core Team. 2005: R: A language and environment for statistical computing. — R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0, Mode of access: http://www.R-project.org.
Rejmanek M., Rejmankova E. Biogeography of artificial islands: effects of age, area, elevation, and isolation on plant species richness // Preslia. 2002. Vol. 74. P. 307–314.
Scherbakov A.V. Investigation and analysis of regional aquatic floras // Hydrobotany: methodology and methods. Rybinsk. 2003. P. 56–69. [In Russian]
Shipunov A.B., Abramova L.A. Floristic changes of the Kem-Ludy islands (1962–2004) // Bull. MOIP, dpt. biol. 2006. Vol. 111. N 1. P. 45-56 [In Russian].
Soberon J.M., Llorente J.B. The use of species accumulation functions for the prediction of species richness // Conservation Biology. 1993. Vol. 7. N 3. P. 480–488.
White sea expeditions of Moscow South-West High School. 2001. Vol. 1. [Electronic resource]. Mode of access: http://herba.msu.ru/belomor/index-en.htm
Vidal E., Madail F., Tatoni Th., Vidal P., Roche Ph. Functional analysis of the newly established plants introduced by nesting gulls in Riou archipelago (Marseille, France) // Acta Oecologica. Vol 19. N. 3. 1998. P. 241–250.
Investigation of the islands of the Kuzokotskaya bay was made during the floristic practice of the Higher plants department (Moscow state university, faculty of biology), lead by Prof. D.D. Sokoloff. Studies of Karelian lakes and islands of the Chupa bay were conducted during White sea expedition of Moscow South-West High School (N 1543), lead by Dr. S.M. Glagolev. Investigation of the lakes in Tver region was carried out on the biological station of the high school “Lake Moldino”. We are grateful to the leaders of floristic groups L.A. Abramova, D.V. Sukhova and E.V. Eliseeva and also to all the members of field research groups.