Лесные экосистемы играют важную роль как в биосфере в целом, так и в жизни людей, являясь важным ресурсом для разных отраслей промышленности (Динамика популяционных генофондов…, 2004). К основным лесообразующим породам относятся представители рода ель (Picea). В Евразии распространены два вида ели – Picea abies (L.) Karst. и P. obovata Ledeb. Первый вид произрастает в Европе, второй – наиболее обычен в Сибири, но доходит до берега Охотского моря. В области соприкосновения ареалов этих видов находится широкая зона гибридизации, которая, вероятно, образовалась в результате их вторичного контакта при отступлении последнего оледенения (Попов, 2003; Попов, Мельник, 2006; Tollefsrud et al., 2008). П.П. Попов (2003) отмечает постепенный морфологический переход между P. abies и P. obovata, поэтому различение видов в зоне гибридизации затруднено. Растения с промежуточными морфологическими признаками часто относят к гибридогенному виду P. × fennica (Regel) Kom.
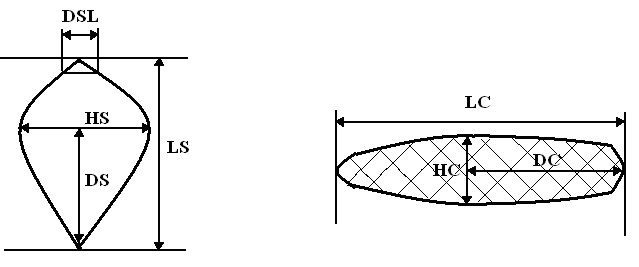
Наиболее удачными диагностическими признаками считают форму семенных чешуй и длину шишек (Раменская, Андреева, 1982; Лантратова, 1991; Скворцов, 2000; Попов, 2003: табл. 1). Для описания формы семенной чешуи используют разные подходы. Словесные формулировки разнообразны и достаточно сложны для применения на практике (табл. 1). П.П. Попов (Попов, 2003; Попов, Мельник, 2006) для формализации этого признака предлагает характеризовать форму семенных чешуй при помощи соотношений их промеров: коэффициентов сужения (DSL/HS*100, где DSL – ширина чешуи на расстоянии 0.1 от ее наибольшей ширины HS) и вытянутости ((LS-DS)/HS*100, где LS – длина чешуи, DS – расстояние от начала чешуи до наибольшей ширины), см. рис. 1. В качестве основного диагностического признака П.П. Попов предлагает использовать разность коэффициентов сужения и вытянутости: для P. abies это значение меньше –5, а для P. obovata – больше 5, причем анализировать необходимо не индивидуальные значения, а среднее для популяции.
Таблица 1. Значения основных диагностических признаков у P. abies и P. obovata
| Вид | Раменская, Андреева, 1982 | Лантратова, 1991 | Скворцов, 2000 | |||
| Форма чешуи | Длина шишки (см) | Форма чешуи | Длина шишки (см) | Форма чешуи | Длина шишки (см) | |
| P. abies | клиновидное основание и неправильно зубчатая верхушка | 10-16 | ромбическая, на верхушке неправильно зубчатая | 10-16 | по верхнему краю остроконечная или тупо треугольная, но не закругленная | 10-15 |
| P. obovata | почковидная, цельнокрайняя | 4-6 (8) | чешуи выпуклые, широкие, вееровидные, на конце цельнокрайние, закругленные | 4-8 | плавно закругленная | 5-8 |
Методика, предложенная П.П. Поповым, снимает трудности, сопряженные с трактовкой словесных описаний формы семенных чешуй, однако довольно трудоемка и плохо применима в полевых условиях. Возможно, форма семенной чешуи хорошо описывается при помощи геометрической морфометрии (метода, позволяющего характеризовать форму как таковую, без использования линейных размеров объекта), однако нам ничего не известно о попытках протестировать эту гипотезу.
Цель нашей работы – выявить морфологический подход, позволяющий эффективно разграничивать P. abies, P. obovata и P. × fennica. В качестве модельной территории мы выбрали подвергавшуюся плейстоценовым оледенениям Северную Карелию, где отмечены все три вида (Кравченко, 2007).
Мы исследовали 141 дерево из 10 популяций ели в мае-августе 2010 года (табл. 2). Наряду с елями из Северной Карелии (80 деревьев из четырех популяций), видовая принадлежность которых не очевидна, мы анализировали P. abies и P. obovata из крайних частей их ареалов (вне зоны гибридизации): три популяции P. abies (из Германии) и три популяции P. obovata (из Ненецкого автономного округа и из Красноярского края). Мы собирали зрелые шишки у деревьев, растущих на расстоянии не менее 10 метров друг от друга. С каждого дерева была собрана одна шишка, за исключением одной популяции P. obovata (табл. 2). Контуры семенных чешуй получали, обводя на бумаге одну чешую из средней части шишки.
Анализ различий по морфологическим признакам мы проводили методами классической морфометрии и геометрической морфометрии.
Мы проводили следующие измерения: длина шишки, наибольшая ее ширина и расстояние до нее от основания шишки; для чешуи делали аналогичные промеры (рис. 1). Вычисляли два вторичных показателя (для шишки и чешуи): отношение длины к ширине и отношение положения наибольшей ширины к длине. Также для семенных чешуй мы высчитывали коэффициенты сужения и вытянутости и их разность (Попов, Мельник, 2006).
Мы анализировали форму семенных чешуй при помощи метода тонких пластин (Павлинов, 2000). Так как нам было затруднительно выбрать точки, гомологичные друг другу на всех контурах, мы использовали 20 меток, расставленных автоматически через равное расстояние по всему контуру чешуи, начиная от ее основания (места прикрепления к шишке). Координаты меток снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig (Rohlf, 2001b). Координаты эталонной конфигурации, значения главных, относительных и частных трансформаций, которые характиризуют меру отличия образца от эталона вычисляли с помощью программы tpsRelw (Rohlf, 2001b). Усредненные контуры чешуй для каждого вида и карельских елей получали при помощи программы tpsSuper (Rohlf, 2003). Редактирование и конвертирование файлов данных мы выполняли во вспомогательной программе tpsUtil (Rohlf, 2000).
Рисунок 1. Схема промеров для семенной чешуи (слева) и шишки (справа)
Для визуализации результатов классической (по совокупности всех промеров) и геометрической морфометрии мы использовали анализ главных компонент. Для оценки достоверности различий морфологических признаков между популяциями использовали однофакторный дисперсионный анализ с последущим попарным сравнением выборок тестом Стьюдента с поправкой Бонферрони. Для статистической обработки данных и построения графиков мы использовали статистическую среду R (R Development Core Team, 2004).
Таблица 2. Характеристики исследованных популяций
| № популяции | Автор сбора | Место сбора | Дата сбора | Число шишек |
| b | Борисова П. | Россия, республика Карелия, Лоухский район, 28 км к СВ от пос. Чупа, южный берег оз. Кривое, около биостанции ЗИН РАН, N 66° 20’ 25’’ E 33° 39’ 20’’ | 20.08.2010 | 20 |
| c | Борисова П. | Россия, республика Карелия, Лоухский район, 29 км к СВ от пос. Чупа, 1,5 км к С-В от биостанции ЗИН РАН, N 66° 20’ 40’’ E 33° 40’ 10’ | 18.07.2010 | 20 |
| d | Борисова П. | Россия, республика Карелия, Лоухский район, 19 км к С-В от пос. Чупа, С-В берег о. Олений, N 66° 18’ 58’’ E 33° 28’ 30’’ | 6.08.2010 | 20 |
| e | Борисова П. | Россия, республика Карелия, Лоухский район, 28 км к СВ от пос. Чупа, северный берег оз. Кривое, N 66° 20’ 51’’ E 33° 38’ 10’’ | 27.07.2010 | 20 |
| f | Абрамова Л. | Россия, Ненецкий автономный округ, 10 км к ЮВ от р. Сямаю, левый берег р. Море-Ю, "Священный лес" | 15.08.2010 | 20 |
| g | Волкова П. | Россия, Красноярский край, юго-западная окраина г. Красноярск, туристическо-экскурсионная зона Государственного Природного Заповедника "Столбы", левый берег ручья Лалетин в его среднем течении N 55° 56’ 48’’ E 92° 44’ 38’’ | 27.08.2010 | 11 |
| h | Волкова П. | Россия, Красноярский край, Туруханский район, западная окраина пос. Бор, левый берег р. Енисей в ее среднем течении, N 61° 36’ 03’’ E 90° 00’ 38’’ | 26.08.2010 | 10 шишек с двух деревьев |
| i | Петров П. | Германия, горный массив Оденваль, окрестности г. Хайдельберг, Gaisberg hill | 1.06.2010 | 18 |
| j | Еськова А. | Германия, окрестности г. Альтлайнинген | 2.05.2010 | 7 |
| k | Петров П., Трушина Е., Трушин К. | Германия, окрестности г. Фрайбург, "Black forest" | 12.05.2010 | 2 |
Результаты классификации исследованных растений по совокупности морфологических промеров представлены на рисунке 2. Видно, что карельские ели не отличаются от «типичных» P. obovata, а «типичные» P. abies характеризуются своеобразной морфологией. Усредненные по популяциям значения разности коэффициентов сужения и вытянутости семенных чешуй для карельских елей превышают +5 (табл. 3), что также характерно, согласно Попову (2003), для типичной P. obovata.
Результаты морфологических промеров и значения коэффициентов сужения и вытянутости, а также их разность (Попов, 2003; Попов, Мельник, 2006) приведены в таблице 3. Типичные P. abies отличаются от карельских елей и типичных P. obovata более крупными и более продолговатыми шишками, а также отрицательными (меньше -5) значениями разности коэффициентов сужения и вытянутости семенных чешуй (табл. 3, рис. 6, 7, эти различия статистически достоверны, p < 0.05).
Достоверные межпопуляционные различия коэффициента вытянутости семенных чешуй почти отсутствуют (только значения для популяции g (P. obovata) достоверно отличаются от значений для популяций i и j (P. abies): p=0.025 и p=0.043 соответственно, табл. 3). Отмечена значительная внутрипопуляционная изменчивость по этому признаку (рис. 3). Межпопуляционные различия коэффициента сужения семенных чешуй выше, значения для популяций «чистых» P. abies и P. obovata не перекрываются, для карельских популяций характерен сильный разброс и промежуточные значения этого признака (табл. 3, рис. 4).
Таблица 3. Значения морфологических промеров для исследованных популяций ели
| Вид | № популяции | LC | LC/HC | DC/LC | LS/HS | DS/LS | CP | CN | CN – CP |
| Карелия, видовая принадлежность не очевидна | b | 62±11 | 1.92±0.30 | 0.39±0.10 | 1.40±0.18 | 0.69±0.05 | 43±7 | 53±11 | 10±14 |
| c | 58±10 | 1.94±0.25 | 0.39±0.09 | 1.31±0.15 | 0.65±0.06 | 45±11 | 56±14 | 10±17 | |
| d | 65±12 | 2.20±0.22 | 0.39±0.12 | 1.35±0.13 | 0.68±0.05 | 43±9 | 58±15 | 15±21 | |
| e | 57±9 | 2.01±0.28 | 0.33±0.1 | 1.36±0.16 | 0.67±0.05 | 45±8 | 59±13 | 15±15 | |
| P. obovata | f | 57±9 | 2.04±0.31 | 0.34±0.08 | 1.32±0.19 | 0.63±0.06 | 50±13 | 71±8 | 21±19 |
| g | 61±8 | 2.17±0.23 | 0.41±0.0 | 1.30±0.11 | 0.71±0.08 | 37±11 | 73±8 | 35±12 | |
| h | 68±4 | 1.99±0.48 | 0.40±0.1 | 1.42±0.08 | 0.68±0.08 | 46±12 | 62±6 | 16±14 | |
| P. abies | i | 118±13 | 2.78±0.23 | 0.40±0.08 | 1.37±0.13 | 0.63±0.05 | 54±20 | 37±8 | -14±13 |
| j | 110±18 | 2.50±0.15 | 0.40±0.07 | 1.48±0.13 | 0.64±0.06 | 53±10 | 43±3 | -10±11 | |
| k | 105±10 | 3.09±0.77 | 0.34±0.03 | 1.37±0.13 | 0.62±0.02 | 52±3 | 40±9 | -12±6 |
Сокращенные обозначения промеров согласно рис. 1. Указано среднее значение ± стандартное отклонение. Приведены также значения коэффициента вытянутости (CP) и коэффициента сужения (CN) и их разности.
Рисунок 2. Распределение исследованных образцов в пространстве двух первых главных компонент (классификация по совокупности морфологических промеров)
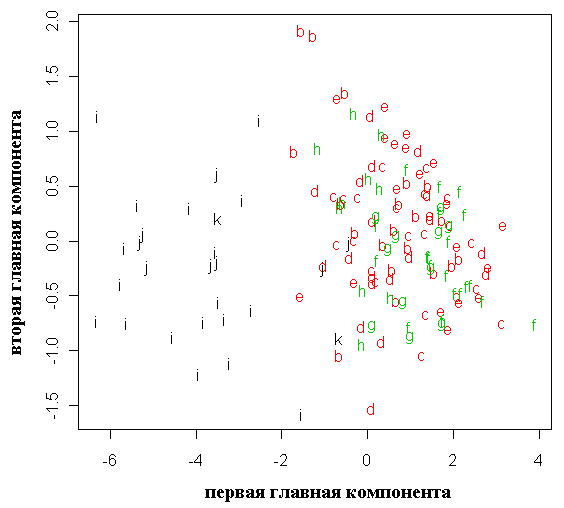
Анализ главных компонент для результатов геометрической морфометрии позволил выделить две отдельные группы образцов по форме семенных чешуй – P. abies и P. obovata; карельские растения образуют промежуточную высоко изменчивую группу, перекрывающуюся с «чистыми» видами, в особенности с последним (рис. 5). Усредненные контуры семенных чешуй для двух видов и карельских елей представлены на рисунке 8, из которого видно, что карельские ели весьма схожи по этому признаку с типичной P. obovata.
Рисунок 3. Значения коэффициента вытянутости для исследованных популяций
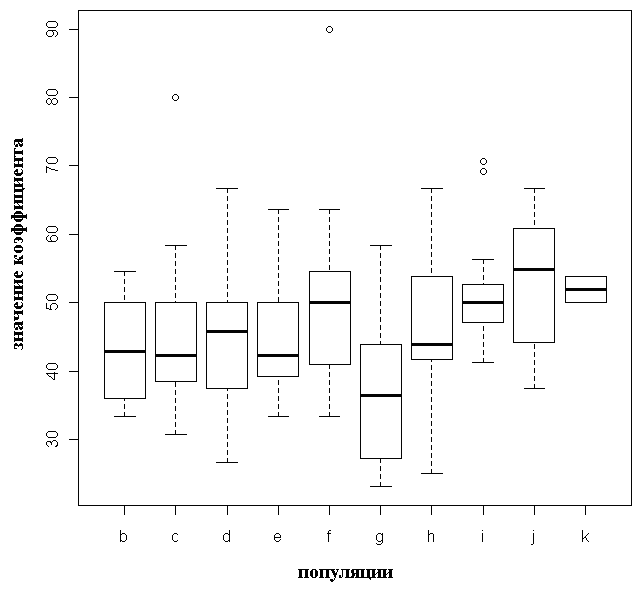
Рисунок 4. Значения коэффициента сужения для исследованных популяций
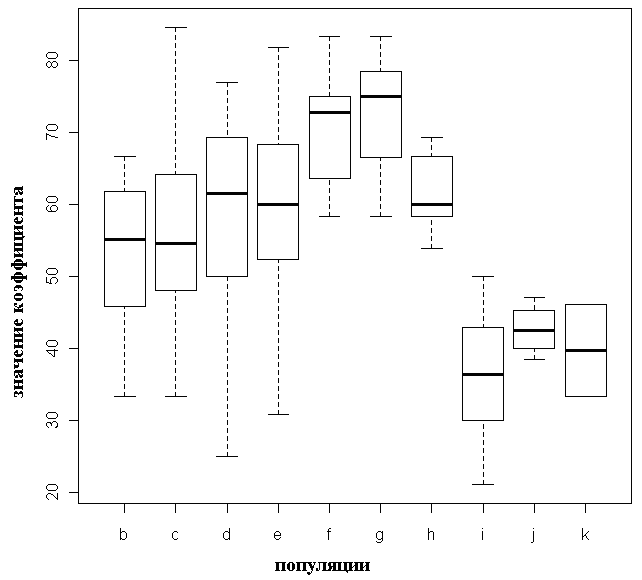
Рисунок 5. Распределение исследованных растений в пространстве двух первых главных компонент для результатов геометрической морфометрии (форма семенных чешуй)
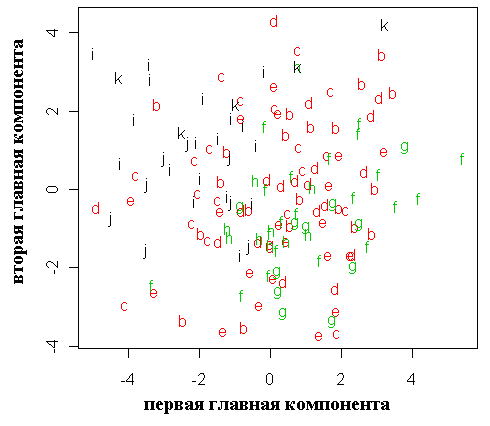
Рисунок 6. Значения разности коэффициентов сужения и вытянутости семенных чешуй для исследованных популяций
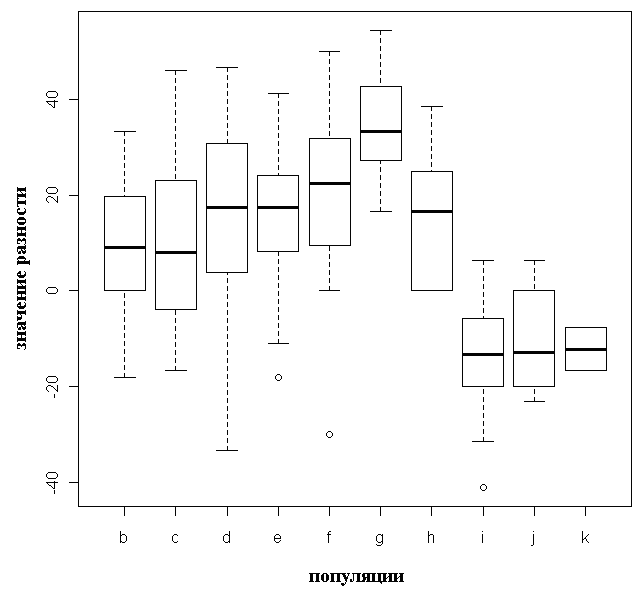
Рисунок 7. Длина шишек для исследованных популяций
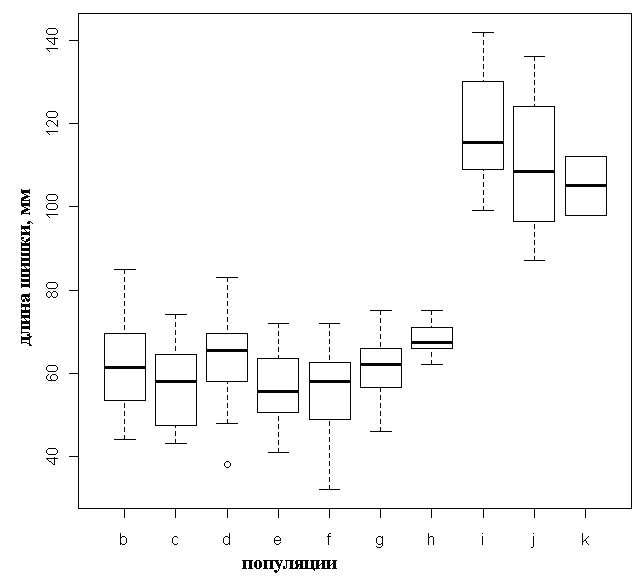
Рисунок 8. Усредненные контуры семенных чешуй

Длина шишек и разность коэффициентов сужения и вытянутости семенных чешуй (Попов, 2003) – удачные диагностические признаки, которые позволяют разграничивать «чистые» P. abies и P. obovata (вне зоны гибридизации). Карельские ели по этим признакам следует относить ко второму виду. В то же время данные геометрической морфометрии свидетельствуют о промежуточной высоко изменчивой форме семенных чешуй карельских елей, что позволяет отнести их к гибридогенному виду P. × fennica.
Таким образом, наши данные допускают две интерпретации. Согласно одной из них, исследованные нами на территории Северной Карелии популяции ели относятся к гибридогенному виду P. × fennica, характеризующемуся промежуточными морфологическими признаками (Раменская, Андреева, 1982). Это выявляется только методами геометрической морфометрии. Согласно другой интерпретации, все карельские популяции ели относятся к P. obovata, на что указывают результаты классической морфометрии (включая коэффициенты, предложенные П.П. Поповым). В таком случае не ясно, почему карельские ели имеют форму чешуй, промежуточную между формами, типичными для двух «чистых» видов.
Для выяснения видовой принадлежности карельских елей необходимо применить методы молекулярно-генетического анализа. Тогда будет ясно, при помощи какого морфологического подхода три симпатрических вида ели могут быть эффективно разграничены в полевых условиях.
Часть материала для настоящей работы собрана в ходе Беломорской экспедиции Московской гимназии на Юго-Западе (№1543). Мы благодарим П. Петрова, Е. Трушину, К. Трушина, А. Еськову и Л. Абрамову за помощь в сборе материала.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |