
Пожары являются одним из основных факторов, влияющих на лесные экосистемы. В последнее время их частота значительно увеличилась в результате антропогенного воздействия [5]. Обычно огонь оказывает на участок леса краткое разрушающее воздействие (за исключением торфяных пожаров, нередко продолжающихся много дней). В результате сгорает лесная подстилка, а при особо сильных низовых пожарах частично разрушается почва.
Изучение пирогенной (послепожарной) сукцессии является актуальной задачей последних лет, так как участившиеся в результате деятельности человека пожары меняют облик лесных экосистем. Необходимо выяснять масштаб и характер этих изменений, вкупе с возможными негативными последствиями.
После пожара начинается восстановление растительного покрова. При послепожарной сукцессии получают возможность развиваться следующие виды растений [1, 4]:
Пирогенная сукцессия представляет собой характерный пример вторичной сукцессии [6]. Как и любая сукцессия, она характеризуется постепенной сменой биоценозов, сопровождающейся увеличением видового богатства (общего числа видов на данной территории) и биомассы населяющих участок организмов. Косвенно об увеличении биомассы растений свидетельствует проективное покрытие, т.е. относительная площадь их проекций на поверхность почвы. Темпы этих процессов замедляются с течением времени, при приближении к климаксной стадии сукцессии.
Еще одно свойство сукцессии – смена жизненных стратегий заселяющихся растений на разных стадиях. В первые годы характерно развитие эксплерентов, отличающихся высокой скоростью размножения и низкой конкурентоспособностью, а также патиентов, приспосабливающихся к неблагоприятным условиям пирогенной экосистемы. Затем они сменяются виолентами, размножающимися медленнее, но вытесняющими другие виды.
Кроме того, на первых стадиях пирогенной сукцессии присутствуют пирофиты – растения, специально приспособившиеся к периодическим пожарам в ходе эволюции [2]. Они используют улучшение условий освещения, обилие элементов минерального питания (интенсивная аммонификация и нитрификация) и резкое снижения конкуренции и быстро осваивают освободившиеся территории. Фитоценозы, содежащие пирофитные растения, часто могут существовать лишь при периодических выгораниях.
Чем на более поздней стадии находится сукцессия, тем более стабильным становится сообщество, так как благодаря большому числу видов энергия используется более полно. К тому же биогенные элементы накапливаются в растениях за счет увеличения их размера [3, 6]. На ранних же стадиях вторичной сукцессии растения еще мелкие, ресурсов достаточное количество, поэтому часто появляются новые виды, большая часть которых через некоторое время вытесняется.
Особого внимания заслуживают участки леса, где в результате продолжительных низовых пожаров выгорела почва. Там обычно не сохраняются семена или подземные части растений, поэтому заселение идет только благодаря диаспорам, появляющимся извне. Кроме того, из-за того, что почва частично разрушена, создаются неблагоприятные условия, нарушающие обычный ход вторичной сукцессии.
Целью данной работы было сравнить ход сукцессии на двух участках выгоревшего соснового леса: том, на котором почва сохранилась и том, где она частично выгорела. Для ее достижения были поставлены задачи: определить видовое богатство, общее проективное покрытие, доминантные виды на площадках с выгоревшей и сохранившейся почвой.
Были выдвинуты следующие гипотезы:
Остров Олений (площадь около 2,7 км2) располагается в губе Чупа Кандалакшского залива Белого моря на территории Керетского природного заказника. В конце июля 2000 года остров горел. Пожар затронул преимущественно северо-восточную часть острова, причем выгорело не менее 30% леса.
Для того чтобы пронаблюдать за ходом пирогенной сукцессии на острове Олений, было заложено две геоботанические площадки размером 10×10 м, на менее (площадка А) и более (площадка Б) поврежденных пожаром участках острова (рис. 1). Площадка А расположена в более сыром месте по сравнению с площадкой Б, что вероятно, и сделало ее менее уязвимой к воздействию огня. Географические координаты центра площадки А: N 66º 18’ 54,6” E 33º 27’ 09,0”. Географические координаты центра площадки Б: N 66º 19’ 00,6” E 33º 27’ 13,1”.
Рисунок 1. Расположение двух пробных площадок на острове Олений
В течение десяти лет (2001-2010 гг.) один раз в год в конце июля – начале августа на обеих площадках оценивалось проективное покрытие высших растений, а также отмечалась степень доминирования видов высших растений в фитоценозе по семибалльной шкале (см. приложение).
Все статистические расчеты и построение графиков проведены в статистической среде R [7]. Для выявления достоверности различий использован статистический тест Вилкоксона. Достоверность связей выявлялась при помощи корреляционного теста Спирмана.
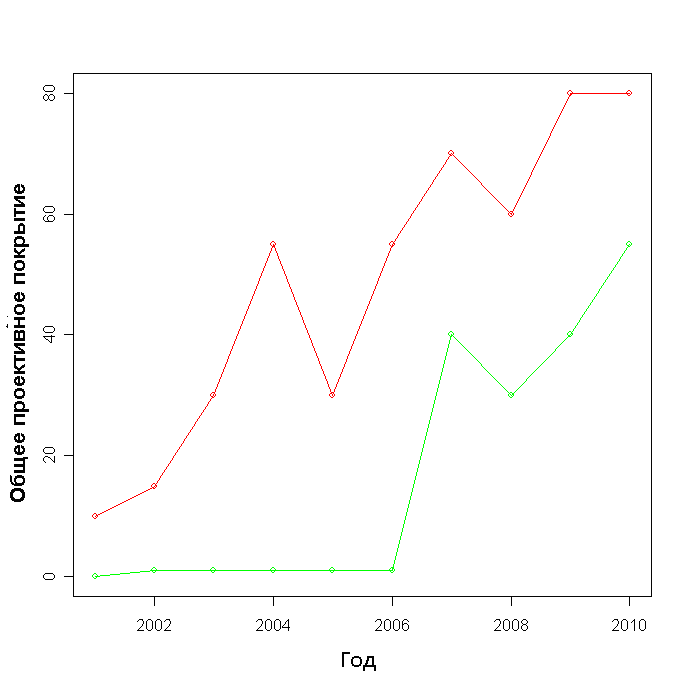
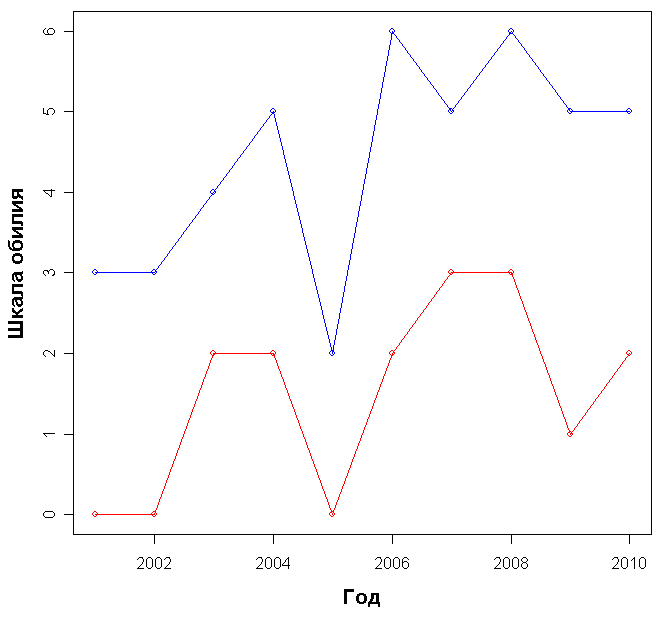
В каждый из годов ОПП на площадке А было больше, чем на площадке Б (рис. 2), различие достоверно (p-value = 0.01). При этом на площадке А наблюдается постоянная тенденция к увеличению ОПП, в то время как на площадке Б она начинает проявляться лишь с 2007 года, а в период с 2001 по 2006 год общий прирост не превышает 1%.
Рисунок 2. Общее проективное покрытие (в %) на площадках
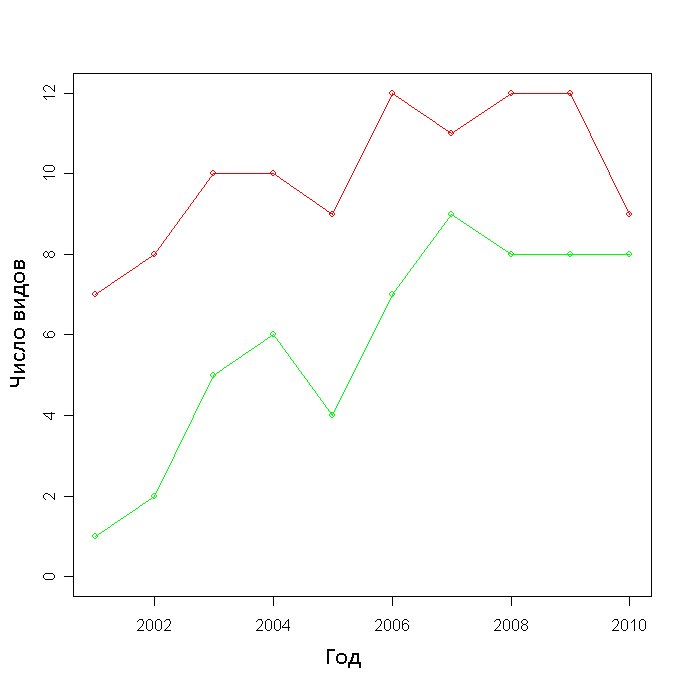
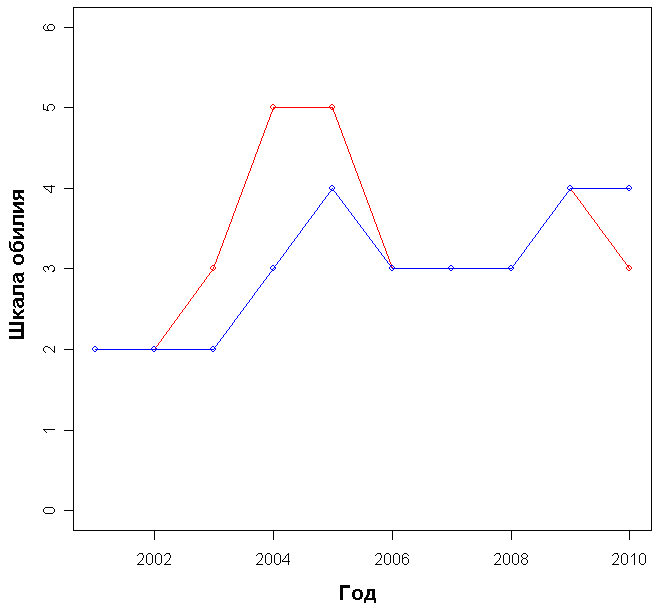
Число видов на участке с выгоревшей почвой было меньше, чем на том, где она сохранилась, во все годы наблюдений (рис. 3). Различие достоверно (p-value=0.002). При этом если на площадке Б число видов увеличивается практически каждый год, то на площадке А нередко наблюдается как увеличение числа видов, так и уменьшение (рис. 3,
см. приложение). Однако их корреляция с годом наблюдения положительна и достаточно достоверна в обоих случаях:
А: r = 0.645, p-value = 0.04
B: r = 0.884, p-value = 0.0007
Рисунок 3. Число видов на площадках
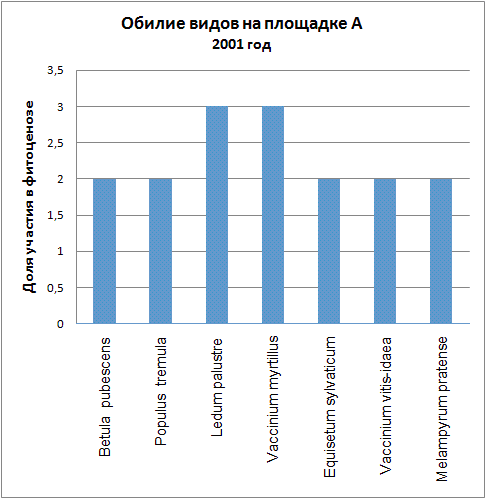
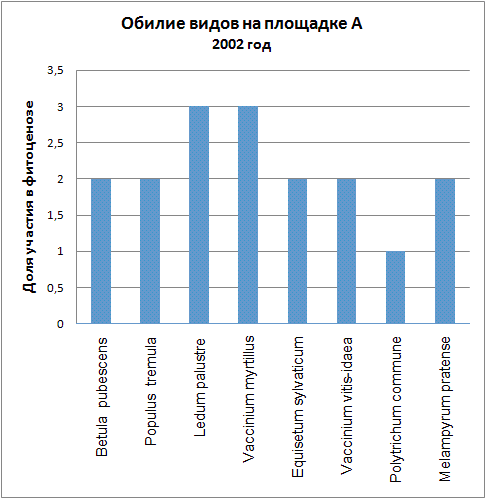
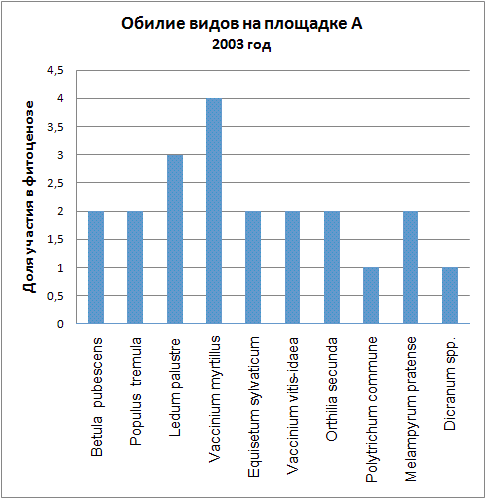
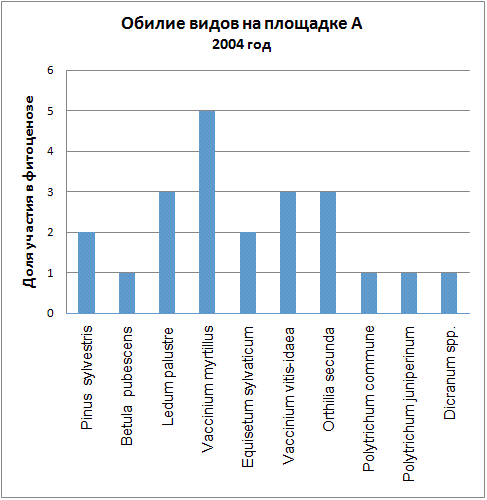
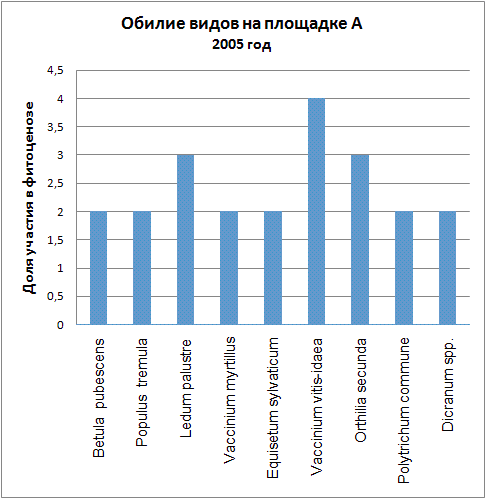
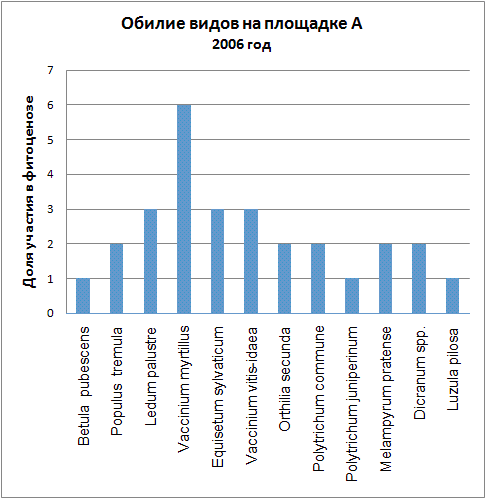
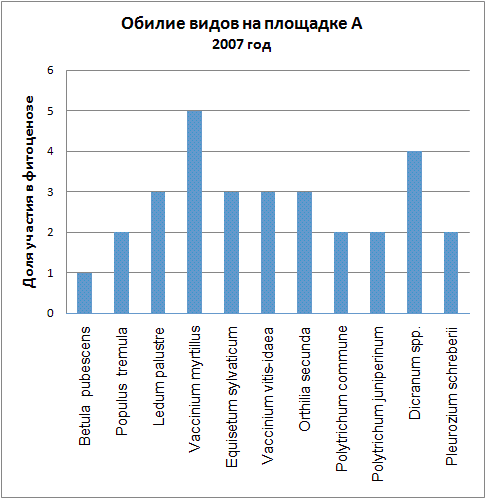
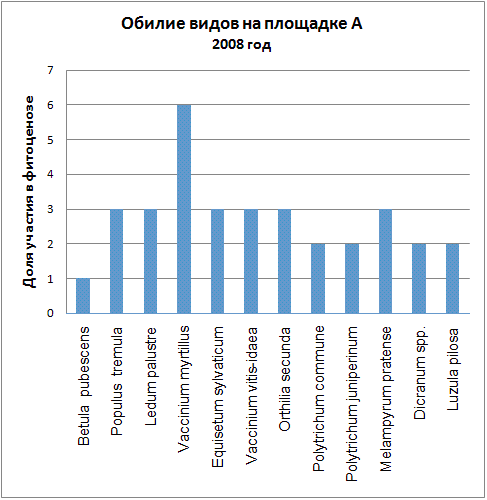
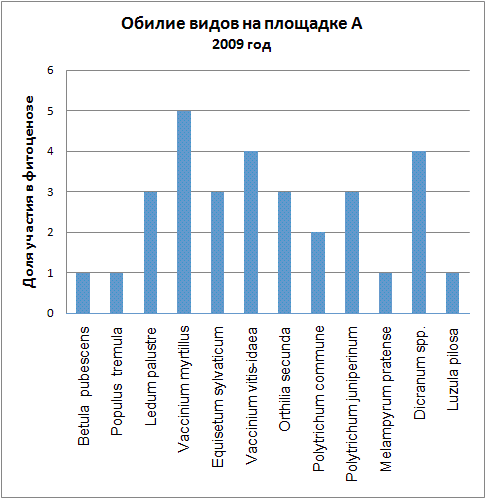
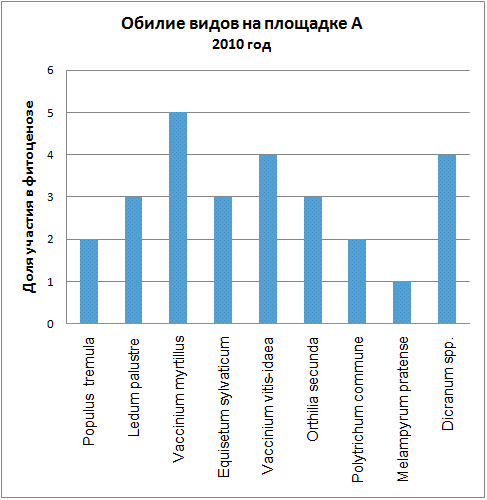
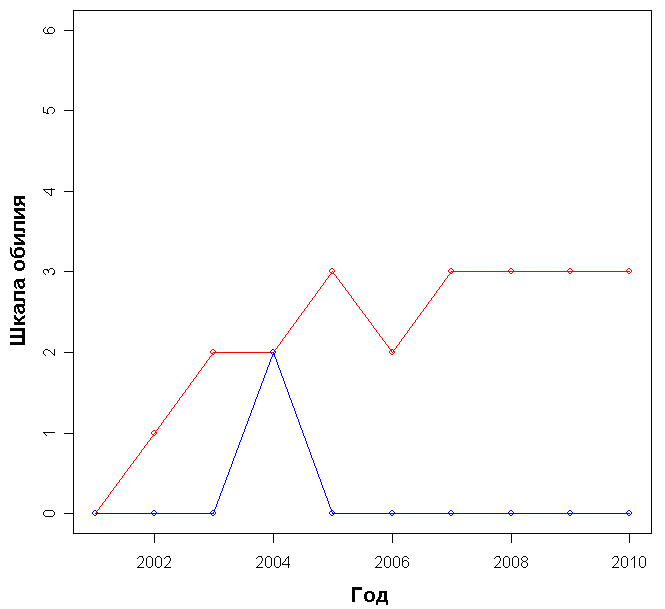
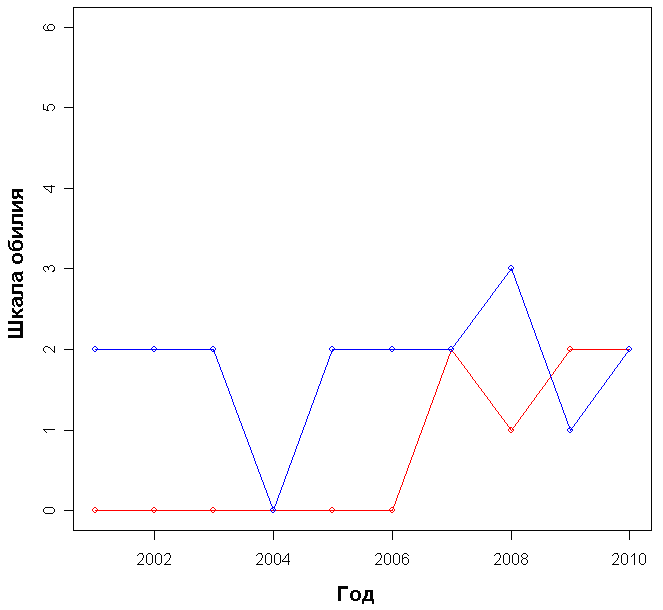
Доминирующими видами считались те, обилие которых превышало значение 4 (их проективное покрытие составляло не менее 25% ОПП). Если таких не было, доминантность определялась по графикам обилия видов в каждый год (см. приложение): выбирали наиболее обильные виды.
Таблица 1. Доминантные виды на площадках в период наблюдения
| Год | Площадка А | Площадка Б |
| 2001 | Ledum palustre, Vaccinium myrtillus | Vaccinium vitis-idaea* |
| 2002 | Ledum palustre, Vaccinium myrtillus | Vaccinium vitis-idaea*, Pinus sylvestris* |
| 2003 | Vaccinium myrtillus | Vaccinium vitis-idaea, Polytrichum juniperinum, Calluna vulgaris |
| 2004 | Vaccinium myrtillus | Vaccinium vitis-idaea, Polytrichum juniperinum, Calluna vulgaris |
| 2005 | Vaccinium vitis-idaea | Vaccinium vitis-idaea |
| 2006 | Vaccinium myrtillus | Polytrichum commune |
| 2007 | Vaccinium myrtillus | Polytrichum juniperinum |
| 2008 | Vaccinium myrtillus | Polytrichum juniperinum |
| 2009 | Vaccinium myrtillus | Polytrichum juniperinum, Calluna vulgaris |
| 2010 | Vaccinium myrtillus | Calluna vulgaris |
*Это единственные виды на площадке в этот год, так что их доминантность носит условный характер
Мы видим, что почти все годы доминантом на площадке А была Vaccinium myrtillus. В первые годы ее кодоминантом являлся Ledum palustre, который затем был вытеснен, а в 2005 году доминантом оказалась Vaccinium vitis-idaea. В итоге можно говорить о двух сменах доминантных видов (2005, 2006) и исчезновении одного кодоминанта (2003).
На площадке Б картина более разнородна. На протяжении двух лет присутствуют три кодоминанта, затем Vaccinium vitis-idaea вытесняет Polytrichum juniperinum и Calluna vulgaris. Еще несколько лет преобладают мхи, а в конце концов доминантом оказывается Calluna vulgaris. Мы наблюдаем исчезновение двух кодоминантов (2005), через несколько лет - появление еще одного (2009) и три смены доминантов (2006, 2007, 2009-10).
Данная работа показывает важность не только предотвращения пожаров, но и их тушения в случае возникновения. Кратковременные пожары лишь возвращают сукцессию на несколько стадий назад, препятствуя возникновению климаксного сообщества, но не разрушая самой экосистемы. Они являются естественным для лесов процессом, благоприятствующим некоторым растениям. В случае же особо сильных пожаров, нарушающих структуру почвы, не только разрушается исходное сообщество, но и замедляется сукцессия в результате отсутствия субстрата для растений.
Видимо, задачей, даже более важной, чем предотвращение пожаров, является их своевременное тушение, которое может предотвратить выгорание почвы и полное разрушение экосистемы.
Я благодарю своих научных руководителей, Полину Андреевну Волкову и Дарью Владимировну Сухову, за руководство и советы; учеников и выпускников гимназии №1543, принявших участие в сборе данных для данных работы; а также Екатерину Слободкину и Евгению Воробьеву, обобщивших материал 2001-2008 годов.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |