Для диагностики видов рода Nymphaea часто используют признаки пыльцевых зерен (Куприянова, 1976; Мейер-Меликян, Диамандопулу, 1996). Существует мнение, что N. candida и N. alba в Европейской России могут быть уверенно разграничены лишь по строению пыльцевых зерен (Куприянова, 1976; Uotilla, 2001). Однако детальные исследования изменчивости этого признака на массовом материале до сих пор не были проведены.
Мы провели промеры пыльцевых зерен кувшинки из восьми регионов Европейской России (42 популяции) под световым микроскопом микроскопом без их предварительной ацетолизной обработки (всего 368 пыльцевых зерен). В качестве показателя вариации признаков мы использовали квартильный размах, так как распределение данных не всегда соответствовало нормальному. Специальное исследование показало, что размеры пыльцевых зерен одного и того же цветка до и после ацетолиза достоверно не различаются, поэтому наши данные сопоставимы с подобными данными (например, Куприянова, 1976), полученными на ацетолизированном материале. Исследование скульптуры экзины было осуществлено при помощи сканирующего электронного микроскопа (далее СЭМ). Для сопоставления с нашими образцами мы исследовали под СЭМ и пыльцу нескольких гербарных образцов из locus typica. Всего было сделано 272 микрофотографии.
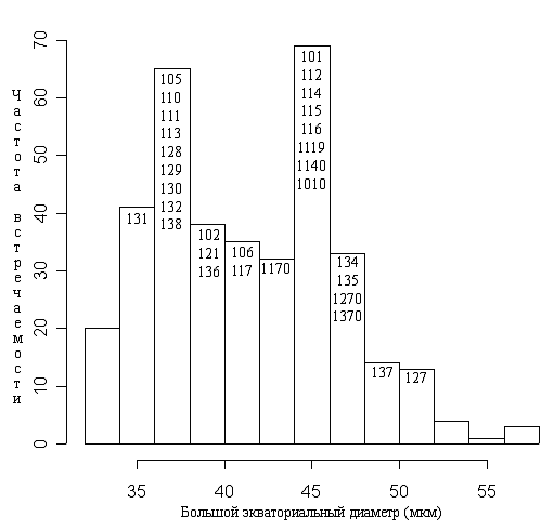
Значения малого и большого экваториальных диаметров пыльцевых зерен имеют бимодальное распределение (рис. 1).
Условные обозначения: числа внутри столбцов диаграммы обозначают номера метапопуляций (согласно табл. 1), медианное значение максимального экваториального диаметра пыльцевых зерен которых совпадает с его значением для данного столбца. Номер метапопуляции с нулем на конце означает другой цветок этой метапопуляции.
Практически все исследованные популяции по этому признаку четко разделяются на две группы: популяции с растениями, имеющими мелкую пыльцу (большой экваториальный диаметр 32-40 мкм, малый – 30-38 мкм), и популяции с растениями, имеющими крупную пыльцу (44-52 мкм и 40-50 мкм соответственно), см. рис. 2. Эти размерные классы соответствуют размерам пыльцевых зерен для N. alba (“мелкая пыльца”) и N. candida (“крупная пыльца”), приведенным Л.А. Куприяновой (1976). Только две исследованные популяции имеют промежуточные размеры пыльцевых зерен (41-46 и 36-43 мкм соответственно). Значения отношения большого экваториального диаметра к малому имеют унимодальное распределение с максимумом на 1,03 и квартильным размахом 1,0-1,1.
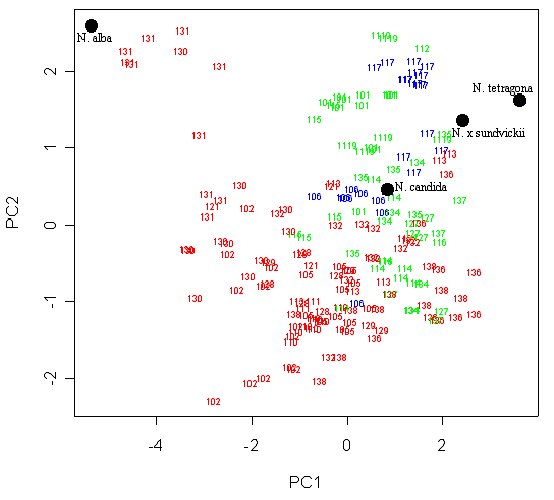
Условные обозначения: число обозначает одно растение, принадлежащее к метапопуляции с соответствующим номером (прил. 1), красным цветом обозначены растения с “мелкими” пыльцевыми зернами, зеленым – с “крупными”, синим – с пыльцевыми зернами “среднего размера”. Черными точками обозначены “якорные образцы”.
Разделение популяций на основании размера пыльцевых зерен соответствует их разделению лишь по совокупности макроморфологических признаков. Это позволило нам сделать предположение о существовании двух хромосомных рас кувшинки на исследованной территории. Это предположение основано на мнении В.А Поддубной-Арнольди (1976) о скоррелированности размера пыльцевых зерен с уровнем плоидности и на данных о большом разбросе подсчитанных чисел хромосом для европейских видов кувшинки (Heslop-Harisson, 1955; Дубына, 1982; Крупкина, 2001).
Визуальный анализ полученных СЭМ-микрографий пыльцевых зерен показал, что скульптура экзины пыльцевых зерен кувшинки неоднородна. Скульптура дистальной части пыльцевого зерна (оперкулума), по нашему мнению, не демонстрирует дискретной изменчивости и представляет собой бугры разного размера, более крупные по краям крышечки, что не согласуется с мнением Л.А. Куприяновой (1976). Скульптура экзины проксимальной части пыльцевых зерен чрезвычайно разнообразна.
Можно выделить пять основных типов скульптуры экзины проксимальной части пыльцевых зерен, между которыми существует непрерывный ряд переходных форм:
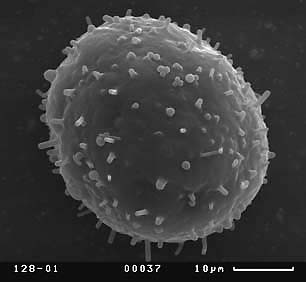



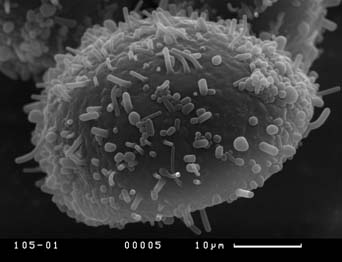
Скульптура экзины нередко существенно различается у разных растений из одной популяции и не коррелирует с размерным классом пыльцевых зерен.
Четвертый тип скульптуры проксимальной части пыльцевых зерен характерен для гербарных образцов N. tetragona из locus typica и отмечен нами у растений из двух популяций с характерными для N. candida значениями макроморфологических признаков. Третий тип скульптуры характерен для гербарных образцов N. candida и N. alba, а также для многих популяций с растениями, обладающих комбинаторными сочетаниями значений макроморфологических признаков, характерных для всех трех видов кувшинки. Таким образом, наши данные не подтверждают мнения Л.А. Куприяновой (1976) о возможности четкого разграничения образцов N. candida и N. alba по скульптуре пыльцевых зерен.
Сканирующая электронная микроскопия, несомненно, позволяет более подробно изучить скульптуру экзины пыльцевых зерен. Обычной является ситуация, когда под световым микроскопом мы видим только палочковидные элементы скульптуры, а на СЭМ-микрографии наряду с ними становятся отчетливо заметны более мелкие бугорчатые или сфероидальные элементы скульптуры. Однако условия высокого вакуума, необходимого для СЭМ, приводят к заметной деформации пыльцевых зерен (рис. 4). Значения малого экваториального диаметра достоверно уменьшаются при СЭМ (t-test: p<<0,01). Значения большого экваториального диаметра у пыльцевых зерен после СЭМ достоверно не изменяются (t-test: p=0,56). Естественно, что при этом на СЭМ фотографиях пыльцевые зерна оказываются гораздо более вытянутыми, чем до напыления в вакууме (большого экваториального диаметра превышает малый в 1,0-1,1 раз до СЭМ и в 1,2-1,5 раз после СЭМ), эти различия высоко достоверны (t-test: p<<0,01).
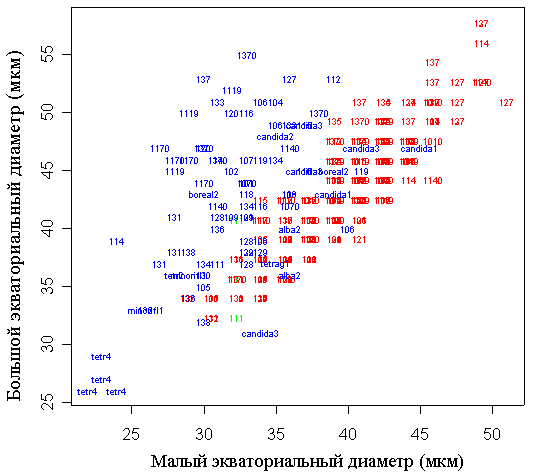
Условные обозначения: числа и число-буквенные комбинации обозначают отдельные пыльцевые зерна, принадлежащие растениям из соответствующих метапопуляций или гербарным образцам, цвет обозначает тип обработки пыльцевых зерен после гербаризации цветков: красный – отсутствие последующей обработки, зеленый – ацетолиз, синий – сканирующая электронная микроскопия
Для оценки таксономической значимости наблюдаемой изменчивости палиноморфологии кувшинки в Европейской России необходимо расширить географию исследований и оценивать уровень плоидности исследуемых популяций.
Дубына Д.В. Кувшинковые Украины. Киев. 1982. 230 с.
Крупкина Л.И. Nymphaeceae Salisb. -- Кувшинковые // Флора Восточной Европы. Т. 10. 2001. С. 25-30.
Куприянова Л.А. Морфология пыльцы видов Nymphaea европейской части СССР // Бот. журн. 1976. Т. 61, № 11. С. 1558-1563.
Мейер-Меликян Н.Р., Диамандопулу Н. Ультраструктура пыльцевых зерен представителей порядка Nymphaeales // Бот. журн. 1996. Т. 81, № 7. С. 1-9.
Поддубная-Арнольди В.А. Цитоэмбриология покрытосеменных растений. М., 1976. 508 с.
Heslop-Harrisson J. Nymphaea // J. Ecol. 1955. Vol. 43. P. 719-734.
Uotila P. Nymphaea L. // Flora Nordica. Vol. 2. 2000. P. 216-221.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |