Проведено исследование морфологии и биологии краснокнижной сапрофитной орхидеи надбородника безлистного (Epipogium aphyllum) на острове Большой Асафьев Кандалакшского залива Белого моря в 2001 – 2005 годах. Наибольшая численность надземной части популяции (151 цветоносный побег) наблюдалась в 2003 году, отдельные месяцы которого были более сухими и теплыми, чем в 2002 и 2004 годах. Наблюдавшиеся изменения численности надземной части популяции и изменения в расположении генеративных побегов в 2003–2004 гг. позволяют предположить либо активное вегетативное или генеративное размножение, либо значительный “запас” нецветущих растений в почве. За 20 часов непрерывных наблюдений за тремя группами цветоносных побегов нами не было зафиксировано ни одного случая взятия насекомыми поллиниев. Возможно, потенциальные опылители надбородника отсутствовали среди насекомых, посетивших это растение во время наблюдений.
Надбородник безлистный (Epipogium aphyllum (F.W. Schmidt) Sw.) – сапрофитная орхидея, ареал которой простирается от центральной Европы на западе до Тайваня на востоке (Chung, Tzeng, 2001) и от Кавказа на юге до Скандинавского полуострова на севере (Leeuwen, 1937). Несмотря на широкий ареал, надбородник считается чрезвычайно редким видом (Красная книга СССР, 1985). Большую часть жизни надбородник существует в виде подземного корневища, на поверхности появляются только цветоносные побеги (Александров и др., 2004). Популяции надбородника в большинстве своём малочисленны и нестабильны. Надбородник, как правило, цветет не ежегодно, и после отцветания растение может не проявлять своего присутствия на протяжение многих лет (Аверьянов, 1988). Наибольшее число генеративных побегов надбородника появляется в относительно тёплые и сухие года (Александров и др., 2004). Цветоносные побеги разлагаются после нескольких недель своего существования (Chung, Tzeng, 2001). Прочие аспекты биологии надбородника остаются неизвестными.
Морфологию надбородника описал достаточно подробно еще в середине XIX века P. Rohrbach (1866). Впоследствии это описание дополнялось публикациями других авторов, из которых особенно значительный вклад внес P. Vermeulen (1965). Некоторые сведения могут быть также подчерпнуты из статей, посвященных близкому тропическому виду надбородника E. roseum (D. Don) Lindley, который отличается от E. aphyllum более крупными размерами (Leeuwen, 1937; Jones, 1985). Размножение надбородника безлистного изучено далеко не окончательно. Считается, что главным образом надбородник размножается вегетативным путём (Красная книга СССР, 1985; Александров и др., 2004). В то же время, по данным некоторых авторов, после образования цветоноса корневище быстро отмирает (Александров и др., 2004). Считается, что цветки надбородника оплодотворяются очень редко, а семена вызревают еще реже (Красная книга СССР, 1985; Александров и др., 2004). P. Rohrbach (1866) считает самоопыление у E. aphyllum невозможным, хотя оно отмечено у близкого вида E. roseum (Jones, 1985). В цветках E. aphyllum содержится большое количество нектара (Кернер фон Марилаун, 1906). Сложно устроенный цветок надбородника требует, по-видимому, специфичных опылителей. P. Rohrbach (1866) и А. Кернер фон Марилаун (1906) видели, как шмель Bombus lucorum (Linnaeus, 1761) посещал цветки надбородника, при этом перенос поллиниев не был отмечен.
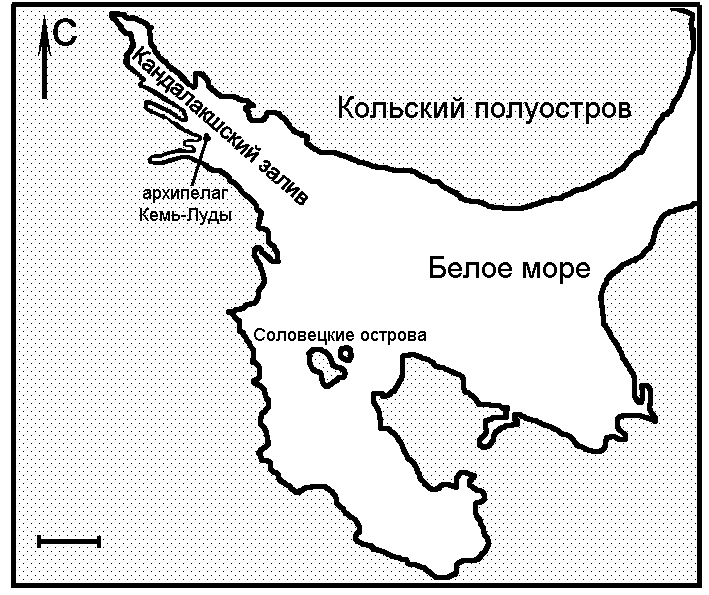
Популяция надбородника на острове Большой Асафьев (иначе называемом Асафий) архипелага Кемь-Луды Кандалакшского залива Белого моря (рис. 1) была впервые обнаружена Н.Е. Богдановой и В.Н. Веховым в 1962 году (Богданова, Вехов, 1969). При повторных поисках популяцию обнаружить не удалось (Воробьёва, Панарин, 1993). В 2001 году надбородник был вновь найден на острове Большой Асафьев участниками Беломорской экспедиции Московской гимназии на Юго-Западе (Сухов, 2001). Эта популяция отличается от других известных нам своей стабильностью (растения цвели ежегодно с 2001 по 2005 годы) и большой численностью (Волкова и др., 2004). Это послужило причиной выбора данной популяции для исследования неизвестных до сих пор аспектов биологии и морфологии надбородника безлистного.
Рисунок 1. Местоположение района исследований
Длина масштабного отрезка 30 км.
В течение четырёх лет (с 2001 по 2004 год) мы подсчитывали число цветоносных побегов в популяции. В 2003 и 2004 годах растения надбородника были закартированы. Картирование проводили методом измерения азимута от растения на два репера на местности. Этот метод позволяет свести к минимуму повреждения растительного покрова при картировании и нанести растения на схему с ошибкой, не превышающей 10% расстояния от растения до репера (Говорухин и др., 1980); то есть не более двух метров в нашем случае. Побеги отмечались на схеме отдельно, если расстояние между ними превышало один метр.
В первой декаде августа 2005 года мы измерили высоту 50 случайно выбранных цветоносных побегов и подсчитали число цветков на них. Мы также отмечали виды сосудистых растений, произраставших в непосредственной близости (на расстоянии менее 10 см) от измеренных цветоносных побегов. Кроме того, было составлено морфологическое описание надбородника безлистного. В это же время (с 9 по 11 августа) были организованы наблюдения за несколькими цветущими растениями для выявления потенциальных опылителей. Мы наблюдали за одиночным цветоносным побегом и за двумя группами побегов различной плотности (3 шт/м2 и 9 шт/м2) с расстояния 1-1,5 метра от соцветия. Всего мы провели 5 сеансов непрерывных наблюдений общей продолжительностью 20 часов: два сеанса в первой половине дня (с 9:30 до 13:30), два сеанса во второй половине дня (с 15:30 до 20:30) и один сеанс ночью (с 00:00 до 02:00). Ночью наблюдения проводили с искусственной подсветкой карманным электрическим фонарем. Мы фиксировали время, когда насекомое посетило надбородник, отмечали отряд насекомого, его приблизительный размер и сколько времени насекомое провело на надбороднике. Также мы отмечали, на какое растение перелетело насекомое, после этого мы по возможности отлавливали насекомое с помощью сачка и определяли его более подробно по определителям (Горностаев, 1999; Определитель насекомых европейской части СССР, 1948).
В работе были использованы данные о метеорологических условиях за годы наблюдений по метеостанции “Кандалакша” (Архив погодных условий, 2005). Статистическая обработка данных была проведена в статистической среде R (R Development Core Team, 2004).
Описание составлено как по литературным данным, так и по данным наших наблюдений в 2001–2005 годах.
Цветоносные побеги надбородника безлистного имеют высоту (5)10–20(30) см (Красная книга СССР, 1985; Chung, Tzeng, 2001; Александров и др., 2004). Растение полностью лишено хлорофилла. Стебель цветоносных побегов безлистный полупрозрачный, палевого цвета. На стебле заметны несколько чешуевидных влагалищ редуцированных листьев. По нашим наблюдениям, подземная часть растения расположена в пределах разлагающегося опада лиственных пород деревьев. Корневище разветвлено в одной плоскости, при этом его толщина примерно равна толщине надземной части стебля. В корневище есть клетки с микоризными грибами (Leeuwen, 1937; Jones, 1985). Известно, что другие бесхлорофилльные орхидеи (например, Corallorhiza trifida), используют подобные грибы для того, чтобы получать органику от фотосинтезирующих растений (McKendrick et al., 2000), но в отношении надбородника таких исследований не производилось.
Цветки надбородника довольно крупные (до полутора сантиметров от кончика гиностемия до цветоножки), располагаются по (1)2–4(8) в соцветии (Красная книга СССР, 1985; Александров и др., 2004). По нашим наблюдениям, высота цветоносных побегов варьирует от 5,5 до 30 см (в среднем 16 см); число цветков на цветоносном побеге колеблется от одного до четырех, чаще всего встречается по два цветка на побеге. Цветоножка изогнута, поэтому цветки направлены вниз. В экспериментах А. Кернера фон Марилауна (1906) цветки после того, как их переворачивали на 180 градусов, в течение 24 часов возвращались в нормальное положение. Губа цветка трёхлопастная, направлена вверх, довольно крупная и окрашена в светло-розовые тона с пурпурным точечным рисунком, на ней виден жидкостный блеск (рис. 2). На краевых участках внутренней стороны поперечного среза губы видны несколько разветвленных выростов, вершины которых составлены клетками с секреторной активностью (Rohrbach, 1866). P. Rohrbach, описавший эти клетки, затруднялся назвать выделяемые ими вещества. Для уверенного определения химической природы этих веществ необходимо провести гистохимические реакции in vivo. По отсутствию достаточно мощных проводящих пучков вблизи от выростов и рыхлой ткани, слагающей губу, можно предположить, что эти клетки выделяют слизеподобные вещества (М.Р. Леонтьева, личное сообщ.). Остальные пять лепестков желтоватые и узколанцетные; имеют одну жилку (Chung, Tzeng, 2001). Эти лепестки располагаются по бокам и в нижней части цветка. Гиностемий слегка изогнут. На нём так же, как и на губе, заметен жидкостный блеск. На верхней стороне гиностемия возле самого рыльца имеется светлое пятно — клеящее основание ножки поллиниев (Кернер фон Марилаун, 1906).
Рисунок 2. Цветок надбородника безлистного
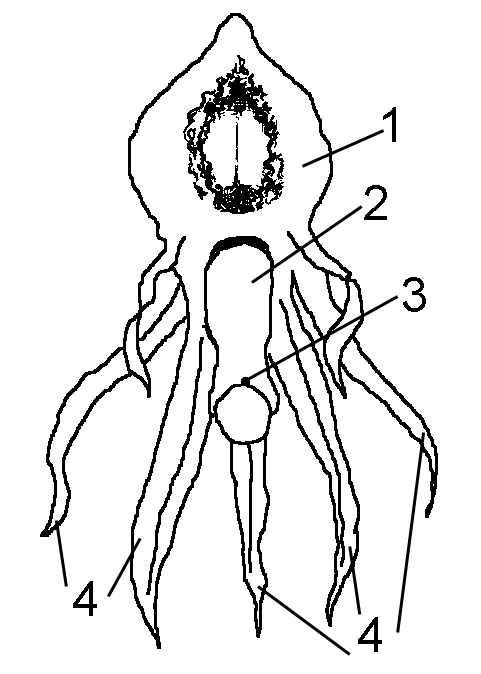
1 -- губа; 2 -- гиностемий; 3 -- клейкое основание поллиниев; 4 -- другие лепестки
Исследуемая популяция надбородника находилась в сыром разнотравном березняке с осиной, примесью рябины и кустарниковых ив с преобладанием в травяном ярусе Athyrium filix-femina, Cirsium heterophyllum и Geranium sylvaticum. В непосредственной близости от цветоносных побегов надбородника обычно произрастали Gymnocarpium dryopteris, Filipendula ulmaria, Angelica sylvestris, Rubus saxatilis и Geranium sylvaticum.Численность надземной части популяции увеличилась в 2002 году по сравнению с 2001 годом и достигла максимума в 2003 году (табл. 1, рис. 3). В 2004 году (по сравнению с 2003 годом) наблюдалось небольшое уменьшение общего числа генеративных побегов (табл. 1, рис. 3) на фоне расширения площади надземной части популяции в северо-восточном направлении (рис. 4). При этом примерная площадь надземной части популяции увеличилась вдвое (с 50 м2 до 100 м2). Повторное картирование надземной части популяции показало также, что местоположение генеративных побегов в 2003 году и в 2004 году различно (рис. 4).
Рисунок 3. Изменение числа генеративных побегов надбородника по годам
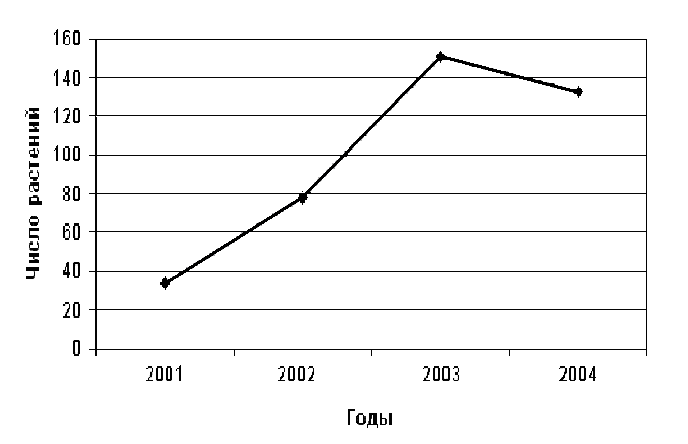
Таблица 1. Численность надземной части популяции надбородника в 2003 и 2004 годах
|
Номер группы (согласно рис. 4) |
Число генеративных побегов в группе |
|
|
2003 год |
2004 год |
|
|
1 |
10 |
12 |
|
2 |
4 |
19 |
|
3 |
2 |
2 |
|
4 |
63 |
14 |
|
5 |
1 |
12 |
|
6 |
1 |
48 |
|
7 |
70 |
6 |
Рисунок 4. Картосхема надземной части популяции Epipogium aphyllum на острове Большой Асафьев архипелага Кемь-Луды
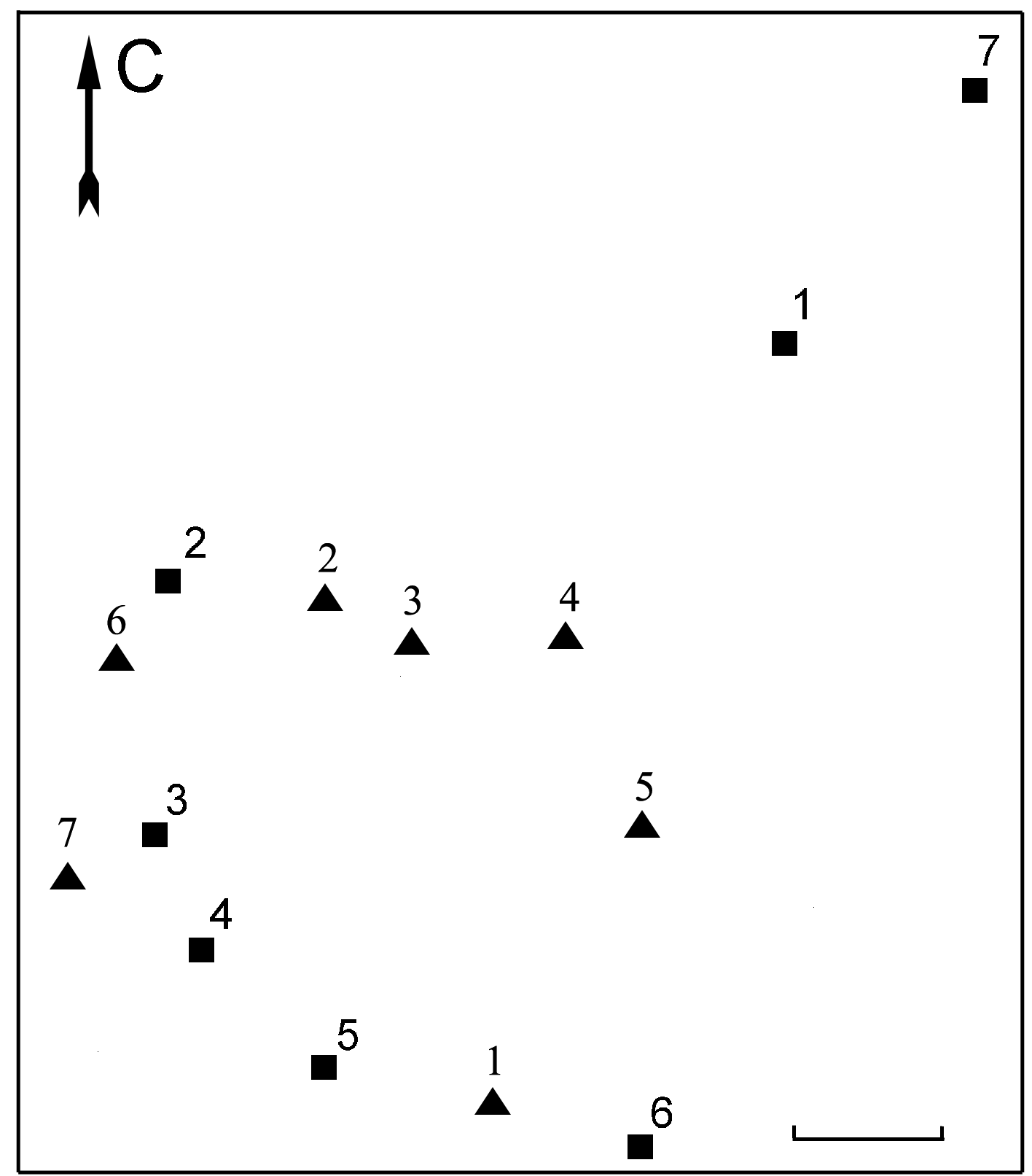
Условные обозначения: Длина масштабного отрезка – 2 м. Форма символа, обозначающего группу цветоносных побегов, соответствует году картирования: треугольник -- 2003 год, квадрат -- 2004 год. Нумерация групп цветоносных побегов соответствует табл. 1.
Анализ метеорологических данных (Архив погодных условий, 2005) показал, что 2003 год отличался от 2002 и 2004 годов более высокими средними температурами марта, мая и августа, а также меньшими значениями относительной влажности воздуха в апреле и июне (рис. 5).
Рисунок 5. Изменение погодных условий в течение года
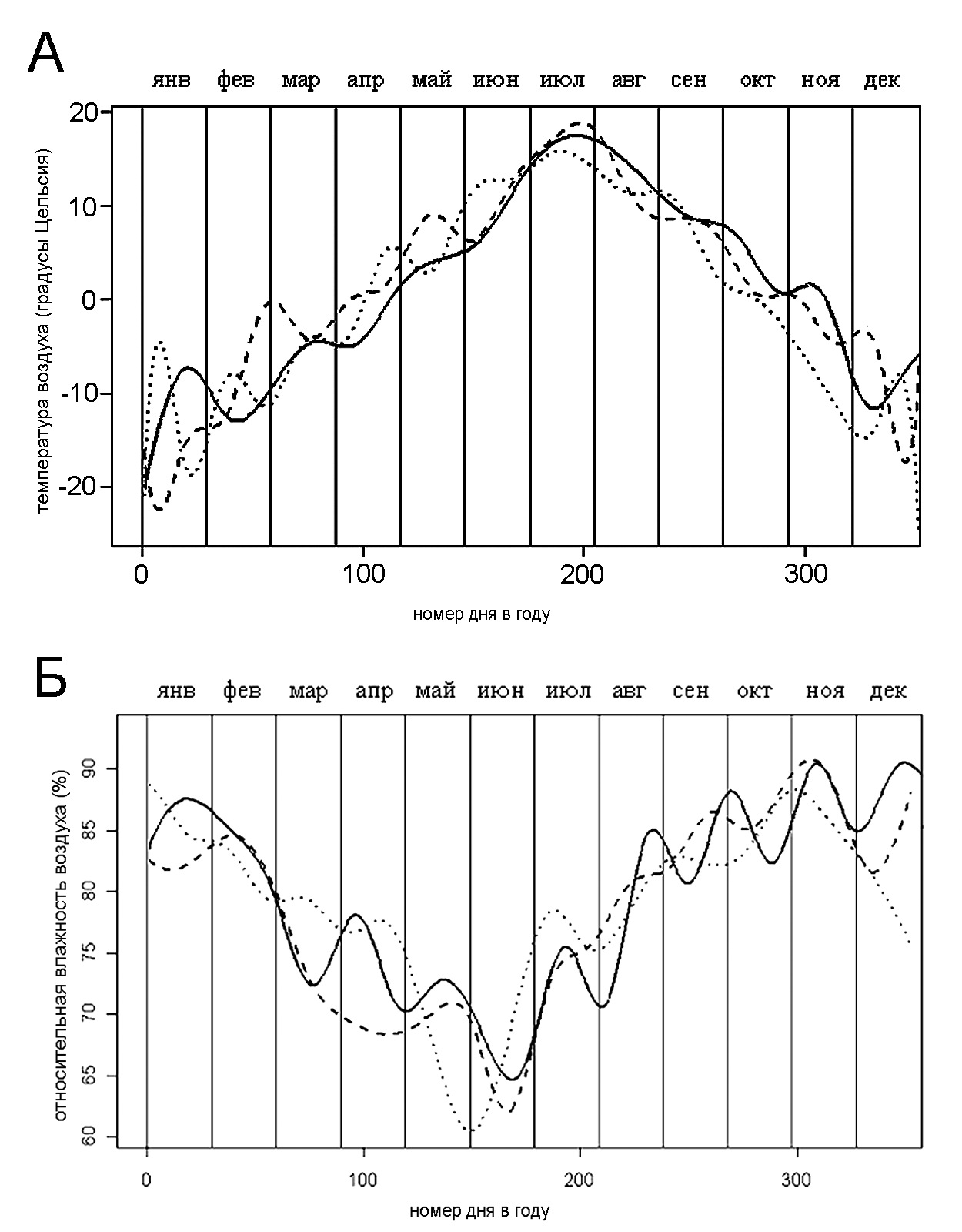
Условные обозначения: Вертикальные линии обозначают первое число месяца. Разные типы линий соответствуют разным годам: короткий пунктир – 2002 год, длинный пунктир – 2003 год, сплошная линия – 2004 год. А – температура воздуха; Б – относительная влажность воздуха
По нашим наблюдениям, цветки посещались исключительно двукрылыми насекомыми (табл. 2). В большинстве случаев (60% от общего числа посетивших цветки насекомых) надбородник посещали насекомые около одного сантиметра длиной. Остальные 40% пришлись на насекомых полусантиметрового размера. Обычно насекомые проводили на надбороднике менее 10 секунд. Иногда насекомое оставалось на растении до 2 минут и более (рис. 6а). Интенсивность посещения надбородника насекомыми зависит от плотности расположения генеративных побегов. Чаще всего посещалась плотная группа побегов (63% от общего числа посещений), чуть реже — диффузное скопление побегов (37%), а одиночный побег посещался в наименьшем числе случаев (10%). Число посещений надбородника насекомыми в первую половину дня и во вторую половину дня примерно одинаково (53% и 47% от общего числа посещений соответственно). Ночью посещений зафиксировано не было.
Таблица 2. Частота посещения цветков надбородника насекомыми разных семейств отряда Diptera
|
Семейство |
Частота посещений: число посещений (доля от общего числа посещений, %) |
|
|
Syrphidae |
Baccha elongata (Fabricius, 1775) |
2 (4) |
|
Cheilosia sp. |
1 (2) |
|
|
Не определено до рода |
14 (24) |
|
|
Anthomyiidae |
3 (5) |
|
|
Muscidae |
3 (5) |
|
|
Lauxaniidae |
самка Meiosimyza decipiens (Loew, 1847), или M. laeta (Zetterstedt, 1847) |
2 (4) |
|
Culicidae |
1 (2) |
|
|
Sphaeroceridae |
1 (2) |
|
|
Не определено до семейства |
30 (52) |
|
Посещая надбородник, насекомые обычно садятся на ярко окрашенную и блестящую сторону губы и активно ползают по ней, “слизывая” с неё нектар, реже зависают в воздухе перед цветком или садятся на стебель и ползают по нему. Насекомые, севшие на цветок, взлетают чаще с гиностемия, сползая на него с губы, или реже взлетают с самой губы (рис. 2).
После посещения надбородника насекомые в основном перелетали на Angelica sylvestris, Geranium sylvaticum, Filipendula ulmaria и Athyrium filix-femina (рис. 6б). Обработка полученных данных (тест хи-квадрат) показала, что насекомые перелетают с надбородника на соседние растения случайно (p=0,15).
Рисунок 6. Продолжительность нахождения насекомых на цветке надбородника и места их вторичной посадки
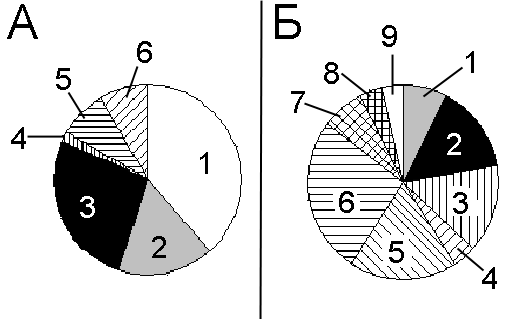
А – продолжительность нахождения насекомых на цветке: 1 -- менее 10 секунд (38% от общего числа посещений); 2 -- от 10 до 30 секунд (17%); 3 -- от 30 секунд до 1 минуты (27%); 4 -- от 1 до 1,5 минут (2%); 5 -- от 1,5 до 2 минут (8%); 6 -- более 2 минут (8%)
Б – места вторичной посадки насекомых после посещения цветка надбородника: 1 -- Rubus saxatilis; 2 -- Athyrium filix-femina; 3 -- Filipendula ulmaria; 4 -- Epipogium aphyllum; 5 -- Geranium sylvaticum; 6 -- Angelica sylvestris; 7 -- Populus tremola; 8 -- Millium effusum; 9 -- Heracleum sibiricum
Результаты морфометрии надбородника безлистного согласуются с литературными данными (Красная книга СССР, 1985; Chung, Tzeng, 2001; Александров и др., 2004), а в некоторых случаях позволяют их дополнить.
Колебания численности надземной части исследуемой популяции находятся, по нашему мнению, в рамках естественных пределов и свидетельствуют о ее хорошем состоянии. Наибольшая численность генеративных побегов наблюдалась в 2003 году, отдельные месяцы которого были более сухими и теплыми, чем в 2002 и 2004 годах. Это подтверждает литературные данные о связи погодных условий и численности надземной части популяций надбородника (Александров и др., 2004). Наблюдавшиеся изменения численности генеративных побегов позволяют предположить либо активное вегетативное или генеративное размножение, либо значительный “запас” нецветущих растений в почве. О подобном “запасе” говорят также изменения в расположении генеративных побегов в 2003-2004 гг.
Наши наблюдения показали, что цветки надбородника имеют нектар, что подтверждает данные А. Кернера фон Марилауна (1906). Этот факт говорит в пользу опыления насекомыми и согласуется с мнением, которое ранее высказал P. Rohrbach (1866). Наши собственные эксперименты показали, что поллинии могут быть отделены от цветка только при движении инородного тела сверху вниз. Такое движение насекомые могли бы совершить, переходя на гиностемий с губы. В то же время нами не было зафиксировано ни одного случая взятия насекомыми поллиниев. Это, вероятно, связано с тем, что насекомые улетали, не достигая клеящего основания поллиниев на гиностемии. Днем в пределах популяции надбородника летало множество шмелей, но ни один из них не пытался сесть на цветок надбородника. P. Rohrbach (1866), напротив, наблюдал посещение надбородника шмелями. Возможно, потенциальные опылители надбородника отсутствовали среди насекомых, посетивших это растение во время наших наблюдений. Это предположение подтверждается и случайным выбором вида растения, на которое перелетали насекомые после посещения надбородника. Интересным представляется наличие завязавшихся семян во вскрытой нами в начале августа 2004 года завязи надбородника, в связи с чем необходимо предположить или наличие опылителей в районе исследований, или возможность самоопыления надбородника. Вопрос о вызревании семян и их прорастании по-прежнему остается открытым.
Наблюдения за посещением надбородника насекомыми позволяют предположить, что эта орхидея использует визуальный или химический способы аттрактации. В пользу этого свидетельствуют связь частоты посещения надбородника насекомыми с плотностью произрастания цветоносных побегов и посещение надбородника насекомыми только в светлое время суток.
Полевые исследования проводились в рамках Беломорской экспедиции Московской гимназии на Юго-Западе (N 1543), проходившей под руководством С.М. Глаголева. Исследования проходили в рамках договора о сотрудничестве с Кандалакшским государственным природным заповедником, заключенного при содействии заместителя директора по научно-исследовательской работе А.С. Корякина. Мы благодарим А.Н. Квашенко за рисунок цветка надбородника, Е.В. Елисееву и Д.А. Чудаева за помощь на полевом этапе исследования, С.А. Брискер и Е.А. Костереву за помощь в работе с собранным материалом по насекомым, Г.В. Фарафонову и А.И. Шаталкина за помощь в определении насекомых, К.А. Астафьева за изготовление срезов губы цветка, В.P. Филина и М.Р. Леонтьеву за консультации по морфологии и анатомии объекта исследований, Т.Н. Виноградову за продуктивное обсуждение работы, а также В.Я. Бергера и В.И. Люляева за помощь в решении транспортных проблем.
Александров Г., Зайцева И., Кобяков К. Редкие растения Мурманской области. Апатиты. 2004. 162 с.
Аверьянов Л.В. Надбородник безлистный // Красная книга РСФСР. Растения. М. 1988. 591 с.
Архив погодных условий [электронный ресурс]. 2005. Режим доступа: http://meteo.infospace.ru.
Богданова Н.Е., Вехов В.Н. Флора сосудистых растений Кемь-Лудского архипелага // Тр. Кандалакшского гос. зап-ка. 1969. Вып. 7. С. 3–59.
Волкова П.А., Сухов С.В., Сухова Д.В., Шипунов А.Б. Состояние популяции Epipogium aphyllum на острове Большой Асафьев (Кемьлудский архипелаг) // Корякин А.С. (ред.) Летопись природы Кандалакшского заповедника за 2004 год (ежегодный отчет). Кандалакша. Т. 1: (Летопись природы Кандалакшского заповедника). 2004.
Воробьёва Е., Панарин А. Надбородник безлистный и дремлик тёмно-красный -- особо редкие орхидные Кандалакшского заповедника // Растения красных книг в заповедниках России: сборник научных трудов. 1993. С. 38–40.
Говорухин А.М., Куприн А.М., Коваленко А.Н., Гамезо М.В. Справочник по военной топографии. М. 1980. 352 с.
Горностаев Г.Н. Определитель отрядов и семейств насекомых фауны России. М. 1999. 176 с.
Кернер фон Марилаун А. Жизнь растений. 5-е изд. СПб. Т. 2. 1906. 838 с.
Красная книга СССР: редкие и находящиеся под угрозой исчезновения виды животных и растений: в 2 т. Т. 2: Растения, грибы и лишайники / Главная ред. коллегия: А.М. Бородин, А.Г. Банников, В.Е. Соколов и др. М. 1985. 480 с.
Определитель насекомых европейской части СССР / Ред. Тарбинский С.П., Плавильщиков Н.Н. М.; Л. 1948. 1128 с.
Сухов С. Находка Epipogium aphyllum на острове Б. Асафьев // Материалы Беломорской экспедиции. Вып. 1 [Электронный ресурс]. 2001. Режим доступа: http://herba.msu.ru/belomor/flora/ep-find.htm
Chung S.W., Tzeng Y.D. Epipogium aphyllum (F.W. Schmidt) Sw. (Orchidaceae), a new addition to the flora of Taiwan // Taiwan Journal for Science. 2001. Vol. 16, N 4. P. 317–320.
Jones D. The self-pollination of Epipogium roseum (D. Don) Lindley // Orchadian. 1985. Vol. 4, N 8. P. 91–92.
Leeuwen W.M. The biology of Epipogium roseum // Blumea. Suppl. I. 1937. Vol. 29, N 6. P. 57–67.
McKendrick S., Leake J., Read D. Symbiotic germination and development of myco-heterotrophic plants in nature: transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections // New Phytol. 2000. Vol. 145. P. 539–548.
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. 2004.
Rohrbach P. Uber die Bluttenbau und die Befruchtung von Epipogium Gmelini. Gottingen. 1866.
Vermeulen P. The place of Epipogium aphyllum in the system of Orchidales // Acta Bot. Neerl. 1965. Vol. 14, N 2. P. 230–241.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |