Известно, что полное выявление видового богатства какой-либо территории невозможно по ряду причин, хотя этот показатель является базовым для многих моделей структуры сообществ и исследований биоразнообразия (Gotelli, Colwell, 2001). Немногочисленные исследования процесса выявления видового богатства определенных территорий основаны большей частью на зоологическом материале (Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005 и ссылки в них, но см. Miller, Wiegert, 1989; Щербаков, 2003). Вероятно, такая ситуация связана с тем, что для зоологов проблема выявления полного видового богатства более актуальна, чем для ботаников, поскольку зоологические объекты хуже поддаются прямому наблюдению. Однако выявление полной флоры также является нелегкой задачей в основном из-за пребывания видов в ненаблюдаемом состоянии (например, в виде банка семян или подземных органов) в определенные сезоны или в отдельные годы и случайного просмотра видов, находящихся в наблюдаемом состоянии (Щербаков, 2003) и, таким образом, требует более подробного изучения. Процесс выявления полной флоры удобнее изучать на объектах с четкими естественными границами, такими, например, как острова и озера.
Со времени своей публикации равновесная модель островной биогеографии (MacArthur, Wilson, 1967) была протестирована на множестве объектов. Немногочисленные исследования были посвящены и динамике флоры островов. Такие исследования основывались на сравнении флористических списков, составленных для одних и тех же островов в разные годы. Различия во флористических списках, полученных для одного острова в разные годы, трактуются как изменения флоры (Abbot, Black, 1980).
Таким образом, исследователи динамики островных флор фактически отождествляют свои флористические списки, полученные в ходе одного полевого сезона, с полной флорой острова, что с очевидностью неверно (Щербаков, 2003). Одни исследователи вообще не затрагивают этот вопрос (Reimanek, Rejmankova, 2002), другие не осознают проблемы, полагая, что при "тщательных исследованиях" флора острова может быть выявлена полностью (Abbot, Black, 1980), третьи признают возможность просмотра отдельных видов (Vidal et al., 1998; Шипунов, Абрамова, 2006), хотя авторы второй статьи считают, что "этот фактор в нашем случае не имеет серьезного значения".
Интересен вопрос и о том, какие именно виды обычно просматриваются. B. Herloff (2003) показала, что могут просматриваться единичные особи, произрастающие в труднодоступных местах; по мнению А. Шипунова и Л. Абрамовой (2006), возможно составить список "наиболее ускользающих видов", который "не имеет почти ничего общего" со списками появляющихся или исчезающих на островах видов. Все это позволяет полагать, что проблема просмотра видов при флористических исследованиях островов в большей степени надумана. Однако показано, что изменения флоры островов идут за счет видов с небольшим обилием (Шипунов, Абрамова, 2006), которые имеют большую вероятность быть незамеченными (Herloff, 2003). Для нас остается неясным, например, какая доля "обнаруженных" на острове видов с малой численностью была просмотрена в ходе предыдущего исследования.
Эти соображения привели нас к необходимости изучения процесса выявления видов растений, находящихся в наблюдаемом состоянии, на двух разных типах территорий с четкими естественными границами (островов и озер) двумя методами, отвечающими специфике избранных объектов (маршрутным методом и методом заложения пробных площадок соответственно).
Мы исследовали четыре острова, расположенных в Кандалакшском заливе Белого моря. Два острова Кузокоцкой губы (Покормежный и Кокоиха) исследованы в начале июля 2003 года, еще два острова губы Чупа (Утичий и Чернышов) исследованы в конце июля 2006 года. Каждый остров обследовался двумя независимыми группами флористов. Для полученных пар флористических списков вычислялись четыре коэффициента, используемые А. Шипуновым и Л. Абрамовой (2006) для характеристики изменений островных флор: коэффициент исчезновения (доля исчезнувших видов от общего числа видов на острове), коэффициент появления (доля появившихся видов), коэффициент стабильности (доля постоянных видов, характеризует изменения видового состава) и коэффициент тенденции (разница между двумя первыми коэффициентами, характеризует изменения видового богатства). При вычислении этих коэффициентов параллельные флористические описания трактовались как описания, сделанные в разные годы. Например, "исчезнувшими" видами считались виды, пропущенные одной из групп. Кроме того, флористические списки для двух островов губы Чупа были сопоставлены с аналогичными списками, полученными в начале августа 2001 года (Беломорские экспедиции…, 2001; Абрамова и др., 2003), для выявления изменений, произошедших во флоре островов за эти пять лет. Во время исследования островов мы использовали метод трансект с обходом части побережья. При этом группа исследователей фронтально проходила остров по предполагаемой максимальной длине, затем по берегу, затем по максимальной ширине и опять по берегу до пункта высадки. Наиболее интересные участки острова (например, болота) обследовали более подробно (Шипунов, Абрамова, 2006).
Для оценки степени согласованности одновременных флористических описаний, выполненных независимыми наблюдателями, мы вычислили каппу Коэна (Cohen’s kappa: Petrie, Sabin, 2000) в статистической среде R (R Development Core Team, 2005), используемый R-скрипт размещен по адресу http://herba.msu.ru/shipunov/software/r/r-en.htm.
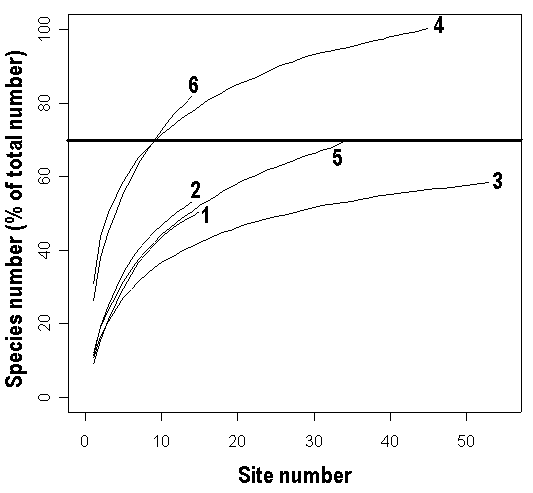
Для оценки скорости выявления полной флоры мы описали флору берегов шести озер (по три в Северной Карелии и в Тверской области) как наиболее флористически богатых типов местообитаний. На берегу каждого озера закладывали ряд пробных площадок и описывали флору каждой из них. Каждая площадка представляла собой участок береговой линии длиной 20 м и имела ширину от начала пояса водной растительности до 1 м от уреза воды. Мы использовали разные алгоритмы размещения пробных площадок. У одного озера (оз. Ближнее) на площадки была разбита вся береговая линия, у другого озера (оз. Молдино) на площадки были разбиты два участка по 500 м на противоположных берегах, на остальных озерах небольшое число (14-34) пробных площадок были равномерно распределены по всей длине береговой линии. Затем для каждого озера были построены «кривые накопления», то есть кривые зависимости общего числа обнаруженных на озере видов от порядкового номера площадки (Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005). Мы использовали усредненные кривые накопления (рис. 1), которые представляли собой результат усреднения 100 кривых, полученных при случайной перестановке порядковых номеров площадок (Gotelli, Colwell, 2001). Для описания кривых накопления наиболее широкое употребление получили три математические модели: экспоненциальная, логарифмическая и модель Кленча. Однако логарифмическая модель не могла быть использована нами для вычисления общего числа видов на озере, поскольку она не имеет предела (Soberon, Llorente, 1993; Diaz-Frances, Soberon, 2005). Полученные нами усредненные кривые накопления лучше описываются моделью Кленча, чем экспоненциальной моделью (сумма квадратов остатков минимальна для модели Кленча). Поэтому общее число видов на озере было вычислено как предел модели Кленча для усредненной кривой накопления видов на этом озере. Мы не смогли воспользоваться программой, предлагаемой для этой цели E. Diaz-Frances и J. Soberon (2005), так как она была недоступна по указанному адресу в Интернет. Поэтому все вычисления и построение графиков были выполнены в статистической среде R (R Development Core Team, 2005).
Значения каппы Коэна составляют 0,7 для островов Кузокоцкой губы и 0,9 для островов Чупинской губы, что свидетельствует о хорошем и отличном согласии между параллельными описаниями соответственно (Petrie, Sabin, 2000).
Доля просмотренных одной из исследовательских групп видов (аналог коэффициентов исчезновения и появления) составила 5-15% от общего числа видов на острове. "Коэффициент тенденции", вычисленный для параллельных описаний одного острова, составлял 0-7%, а "коэффициент стабильности" -- 74-90% (табл. 1).
Таблица 1. Значения используемых коэффициентов для параллельных описаний островов
| Остров | Всего видов | "Коэффициент" | |||
| исчезновения | появления | тенденции | стабильности | ||
| Покормежный | 180 | 0.11 | 0.06 | 0.00 | 0.90 |
| Кокоиха | 131 | 0.05 | 0.05 | 0.05 | 0.83 |
| Утичий | 97 | 0.15 | 0.09 | 0.07 | 0.74 |
| Чернышов | 90 | 0.11 | 0.09 | 0.02 | 0.80 |
Список видов, исчезнувших или появившихся на Кемь-Лудских островах, как справедливо отмечают А. Шипунов и Л. Абрамова (2006) имеет почти ничего общего со списком видов, просмотренных одной из групп на островах Кузокоцкой губы (два совпадающих вида: Carex buxbaumii и Scutellaria galericulata -- 4% от "Кемь-Лудского списка") и со списком видов, просмотренных на островах Чупинской губы (только Melampyrum sylvaticum).
Список видов, появившихся или исчезнувших на островах Чупинской губы, также не имеет почти ничего общего со списком просмотренных видов на островах Кузокоцкой губы (два общих вида: Achillea millefolium и Carex globularis). Зато более половины (52%) появившихся на островах Чупинской губы видов были просмотрены одной из групп (по 6 видов каждой группой).
Просмотренные виды, как правило, не очень редки на острове (чаще, чем "единично"), относятся к разным жизненным формам (деревья -- Alnus incana, кустарники -- Salix caprea, Rosa majalis и др., многочисленные травянистые растения), произрастают в различных биотопах (от сфагновых болот в центральной части острова: Rubus chamaemorus до средней литорали: Zostera marina) и имеют разный внешний вид (от невзрачного Poa pratensis до яркой Coronaria flos-cuculi).
На близко расположенных островах со сходной флорой, как правило, просматривались разные виды. Исключение составляют Botrychium boreale, Corallozhiza trifida, Goodyera repens, Juncus athrofuscus, Lycopodium annotinum, Carex globularis, C. aquatilis и Galeopsis bifida, которые просматривались разными группами на двух-трех островах из Кузокоцкой губы и из губы Чупа (табл. 2).
Таблица 2. Просматриваемые, исчезнувшие и появивившиеся виды (растения расположены по алфавиту)
| Просматриваемые виды | Острова губы Чупа: 2001-2006 гг. | ||
| острова Кузокоцкой губы | острова губы Чупа | исчезнувшие виды | появившиеся виды |
| Achillea millefolium, Agrostis gigantea, Alnus incana, Angelica sylvestris, Botrychium boreale, B. lunaria, Calamagrostis purpurea, Carex buxbaumii, C. globularis, C. lasiocarpa, C. rotundata, Corallorhiza trifida, Dactylorhiza maculata, Deschampsia caespitosa, Equisetum pratense, Galeopsis bifida, Glaux maritima, Goodyera repens, Hippuris tetraphylla, Juncus artrofuscus, Lathyrus palustris, Lycopodium annotinum, Moneses uniflora, Oxycoccus microcarpus, Poa nemoralis, P. glauca, Ranunculus reptabundus, Rosa majalis, Rumex aquaticus, Salix glauca, S. hastata, S. lapponum, Scutellaria galericulata, Selaginella selaginoides, Stellaria crassifolia | Agrostis stolonifera, Antennaria dioica, Calamagrostis neglecta, Carex aquatilis, C. brunescens, C. glareosa, C. globularis, C. mackenziei, C. rariflora, C. rostrata, C. salina, Cerastium holosteoides, Cochlearia groenlandica, Conioselinum tataricum, Coronaria flos-cuculi, Erysimum hieracifolium, Equisetum fluviatile, E. sylvaticum, Galeopsis bifida, Gymnocarpium dryopteris, Heracleum sibiricum, Hieracium spp., Huperzia selago, Lycopodium annotinum, L. complanatum, Melampyrum sylvaticum, Parnassia palustris, Polygonum viviparum, Poa pratensis, Polypodium vulgare, Ranunculus polyanthemos, Rubus chamaemorus, Salix caprea, Sedum acre, Stellaria media, Zostera marina |
Achillea millefolium, Agrostis tenuis, Comarum palustre, Eleocharis uniglumis, Epilobium palustre, Polygonum boreale, Potentilla egedii, Stelaria graminea, Vicia sepium |
Agrostis gigantea, A. stolonifera, A. straminea, Antennaria dioica, Aster tripolium, Campanula rotundifolia, Carex aquatilis, C. glareosa, C. globularis, C. mackenzii, C. salina, Heracleum sibiricum, Huperzia selago, Parnassia palustris, Poa pratensis, Pyrola chlorantha, P. minor, Stellaria media, Vicia cracca, Festuca rubra, Ruppia maritima, Sagina nodosa, Sedum acre, Solidago virgaurea, Triglochin palustre, Zostera marina |
Число пробных площадок, необходимых для выявления флоры берегов озера на рекогносцировочном уровне (все виды со встречаемостью «изредка» и выше, 70% от полной флоры: Щербаков, в печати), практически не зависит от размера озера и от региона (рис. 1, табл. 3). Об этом свидетельствует и заметное уменьшение части береговой линии, которую нужно для этого исследовать, по мере увеличения размеров озера (табл. 3). Таким образом, вне зависимости от размеров озера для рекогносцировочного описания его флоры в среднем необходимо исследовать 9-25 пробных площадок (200-500 м береговой линии).
Таблица 3. Основные характеристики исследованных озер
| Озеро | Длина береговой линии (м) | Исследовано* | Общее число видов** | Обнаружено 70% от общего числа видов* |
| Тверская область | ||||
| Ольшево | 6 350 | 15 (5) | 153 | 19 (6)*** |
| Чеполшево | 5 000 | 14 (6) | 199 | 14 (6) |
| Молдино | 26 200 | 53 (4) | 169 | 18 (1) |
| республика Карелия | ||||
| Ближнее | 900 | 45 (100) | 45 | 11 (24) |
| Долгое | 4 320 | 34 (16) | 166 | 21 (10) |
| Нижнее Пулонгское | 9 540 | 14 (3) | 124 | 9 (2) |
* -- число площадок (длина береговой линии, % от общей длины)
** -- вычислено как предел модели Кленча (Soberon, Llorente, 1993).
*** -- вычислено путем экстраполяции модели Кленча (Soberon, Llorente, 1993).
Рисунок 1. Усредненные кривые накопления для разных озер.
1 – оз. Ольшево, 2 - оз. Чеполшево, 3 - оз. Молдино, 4 - оз. Ближнее, 5 - оз.
Долгое, 6 - оз. Нижнее Пулонгское. Общее число видов на озере, принятое за 100%,
вычислено как предел модели Кленча (Soberon, Llorente, 1993).
Наши исследования процесса выявления флоры озер показали, что даже на рекогносцировочном уровне обнаружение находящихся в наблюдаемом состоянии видов относительно бедных частных северотаежных флор требует значительных усилий. Этот результат подтверждается и большим расхождением (до 15% от общего числа видов) в одновременных независимых описаниях островов.
Одновременные независимые описания островов показали, что виды просматриваются совершенно случайно, вне зависимости от их габитуса и биотопической приуроченности. По этой причине нам кажется необоснованным анализировать появление или исчезновение отдельных видов на острове, так как оно может быть обусловлено просмотром вида во время более раннего исследования (в этом случае будет наблюдаться "появление" вида) или во время более позднего исследования ("исчезновение" вида). С другой стороны, комплексный анализ динамики отдельных видов (при объединении данных по различным островам), как это было сделано А. Шипуновым и Л. Абрамовой (2006), представляется нам удачным подходом. Как было показано нами, на разных (пусть и очень сходных) островах просматриваются разные виды, таким образом, при объединении данных по ряду сходных островов последствия случайного просмотра отдельных видов "взаимоуничтожаются".
Таким образом, флористическое описание можно рассматривать как своего рода выборку из генеральной совокупности, которой является полная флора данной территории. Поэтому сравнение флористических описаний (в частности при выявлении динамики островной флоры) следует проводить, используя логику сравнения статистических выборок. В статистике достоверность различия между выборками устанавливают путем сравнения вероятности существования этого различия с установленным пороговым значением. В исследованиях динамики островных флор вероятность значимых изменений во флоре острова может характеризоваться коэффициентами, описывающими эти изменения (Шипунов, Абрамова, 2006). Пороговыми значениями, вероятно, следует считать значения коэффициентов, полученные при сравнении параллельных описаний одной и то же флоры (аналог двух выборок, полученных из одной генеральной совокупности). Такие описания будут различаться только из-за случайного просмотра отдельных видов каждой из групп. Наши исследования показали, что для коэффициентов появления и исчезновения пороговое значение можно принять равным 0,15, для коэффициента стабильности -- 0,7, а для модуля коэффициента тенденции -- 0,1. Это значит, что если при сравнении двух описаний флоры острова, сделанных в разные годы, значение коэффициента стабильности не превосходит своего порогового значения, а значения остальных коэффициентов не превышают его, то на исследуемом острове произошли соответствующие изменения флоры, отражающие реальную динамику, а не погрешности описания.
Отметим, что использование предлагаемых пороговых значений позволяет выявить реальные (не вызванные случайным просмотром отдельных видов, находящихся во время исследований в наблюдаемом состоянии) изменения островной флоры. Нужно учесть, что эти изменения могут быть связаны с различиями в погодных условиях в годы исследований (в неблагоприятный год некоторые растения могут не вегетировать, находясь в "ненаблюдаемом" состоянии), а не с динамикой островной флоры в рамках равновесной модели островной биогеографии (MacArthur, Wilson, 1967). Вероятно, для выяснения причин наблюдаемого изменения флоры необходимо проводить серию (не менее четырех) флористических описаний острова, выполненных с интервалом в 1-2 года. Однако, помимо большой трудоемкости такого метода, его существенным недостатком является возможное влияние на флору острова деятельности исследователей.
Разумеется, полученные нами закономерности выявления находящихся в наблюдаемом состоянии видов не могут быть механически распространены на все флористические исследования. Следует ожидать, что конкретные параметры этих закономерностей (например, пороговые значения коэффициентов, характеризующих динамику островных флор) будут варьировать в зависимости от многих факторов, таких как размер исследуемой территории, природная зона, квалифицированность исследователей и т.п. К примеру, в наших исследованиях согласованность между одновременными независимыми описаниями флоры островов была выше на островах губы Чупа по сравнению с островами Кузокоцкой губы. С одной стороны, это можно объяснить меньшим размером Чупинских островов. С другой стороны, описания на островах губа Чупа проводили участники Беломорской экспедиции Московской Гимназии на Юго-Западе, имеющие большой опыт совместной деятельности подобного рода, тогда как описания на островах Кузокоцкой губы проводились нескоординированными группами.
Не претендуя на универсальность описанных закономерностей процесса выявления флоры, мы хотели бы подчеркнуть, что подобные исследования необходимы для определения "чувствительности" избранного метода флористических изысканий. Решение конкретных флористических задач (в особенности связанных со сравнением флор разных территорий или в разные моменты времени) без информации о «чувствительности» используемого метода подобно измерениям, выполненным при помощи линейки с неизвестной ценой деления. Такие данные представляют небольшую научную ценность. Оценка доли обнаруженных видов и прогнозирование общего числа видов на исследуемой территории при помощи кривых накопления необходима при проведении инвентаризационных работ, что неоднократно отмечалось рядом исследователей (Miller, Wiegert, 1989; Soberon, Llorente, 1993; Gotelli, Colwell, 2001; Diaz-Frances, Soberon, 2005). Подобная проблема определения чувствительности сложного метода в последнее время стала актуальной в совершенно иной области исследований – непрямом анализе последовательностей ДНК – в которой, как и во флористических исследованиях, человеческий фактор играет заметную роль. Для решения этой проблемы молекулярные биологи настоятельно рекомендуют определять чувствительность метода при каждом исследовании и обязательно указывать ее в публикациях (Bonin et al., 2004). Нам представляется, что флористы должны последовать этому примеру.
Беломорские экспедиции Московской гимназии на Юго-Западе. 2001. Вып. 1. [Электронный ресурс]. Режим доступа: http://herba.msu.ru/belomor
Шипунов А.Б., Абрамова Л.А. Изменения флоры островов Кемь-Лудского архипелага (1962–2004) // Бюлл. МОИП, сер. биол. 2006. Т. 111. N 1. С. 45-56.
Щербаков А.В. Изучение и анализ региональных флор водоемов // Гидроботаника: методология, методы: Материалы школы по гидроботанике (п. Борок, 8—12 апреля 2003 г.). — Рыбинск, 2003. — С. 56—69.
Abbott J., Black R. Changes in species composition in floras on islets near Perth, Western Australia // Journ. Biogeogr. 1980. Vol. 7. N 4. P. 399–410.
Bonin A., Bellemain E., Eidesen P.B., Pompanon F., Brochmann C., Taberlet P. How to track and assess genotyping errors in population genetics studies // Molecular Ecology 2004. Vol. 13. P. 3261-3273.
Diaz-Frances E., Soberon J. Statistical estimation and model selection of species accumulation curves // Conservation Biology. 2005. Vol. 19, N 2. P. 569-573.
Gotelli N.J., Colwell R.C. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness // Ecology Letters. 2001. Vol. 4. P. 379–391.
Herloff B. Floristic changes during the twentieth century in the southern archipelago of Goeteborg, SW Sweden // Svensk Bot. Tiddskr. 2003. Vol. 97. P. 3–14.
McArthur R.H., Wilson O.E. The theory of island biology. Princeton, New Jersey, 1967. 203 p.
Miller R.I., Wiegert R.G. Documenting completeness, species-area relations and the species-abundance distribution of a regional flora // Ecology. 1989. Vol. 70, N 1. P. 16-22.
Petrie A., Sabin C. Medical statistics at a glance. Blackwell science, London. 2000. 138 p.
R Development Core Team. 2005: R: A language and environment for statistical computing. — R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0, Mode of access: http://www.R-project.org.
Rejmanek M., Rejmankova E. Biogeography of artificial islands: effects of age, area, elevation, and isolation on plant species richness // Preslia. 2002. Vol. 74. P. 307–314.
Soberon J.M., Llorente J.B. The use of species accumulation functions for the prediction of species richness // Conservation Biology. 1993. Vol. 7. N 3. P. 480–488.
Vidal E., Madail F., Tatoni Th., Vidal P., Roche Ph. Functional analysis of the newly established plants introduced by nesting gulls in Riou archipelago (Marseille, France) // Acta Oecologica. Vol 19. N. 3. 1998. P. 241–250.
Исследования островов Кузокоцкой губы были сделаны во время флористической практики кафедры высших растений биологического факультета МГУ под руководством Д.Д. Соколова. Исследования островов Чупинской губы и карельских озер были сделаны во время Беломорской экспедиции Московской Гимназии на Юго-Западе (№1543), проходившей под руководством С.М. Глаголева. Исследования озер в Тверской области были проведены на биостанции гимназии «Озеро Молдино». Мы благодарим старших флористических групп Л.А. Абрамову, Д.В. Сухову и Е.В. Елисееву, а также всех, кто участвовал в описании островов и озер.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |