[презентация (PDF-файл)] [2001 год] [2003 год]
Осоки (Carex L.) - крупнейший род семейства осоковые (Cyperaceae), насчитывающий более 1000 видов, многие из которых являются трудноразличимыми. Определение осок осложняется еще и тем, что близкие виды осок способны к гибридизации. Гибридные растения имеют промежуточную морфологию и значительно затрудняют определение. Два, по-видимому гибридных, вида приморских осок Северного полушария - Carex salina и Carex recta - заслуживают особого внимания (Haines, 2000; Standley, 1990; Cayouette, Morisset, 1985).
Большинство гибридов осок являются спорадическими гибридами первого поколения, которые стерильны или имеют пониженную фертильность, однако некоторые становятся стабилизированными и широко распространенными. Термин "стабилизированный гибрид" означает, что таксон имеет гибридную природу, однако ведет себя как особый вид. К чертам, часто ассоциирующимся с стабилизированными гибридами, относятся: относительно однообразная морфология, морфологические отличия от гибридов первого поколения, способность производить жизнеспособное потомство, а также более или менее широкое распространение, включая местонахождения, в которых один или оба родительских вида не встречаются (Haines, 2000).
Cayouette и Morisset (1985) предположили, что C. recta и C. salina в Северной Америке имеют гибридную природу, основываясь на нарушенном мейотическом поведении их хромосом, низкой фертильности пыльцы и небольшой семенной продуктивности. На основе кариологических данных они выдвинули гипотезу, по которой североамериканские C. salina - это более или менее стабилизировавшийся гибрид C. paleacea x C. subspathacea, а североамериканские C. recta - это стабилизировавшийся продукт гибридизации C. paleacea x C. aquatilis. Standley (1990) подтвердил эту гипотезу, используя электрофорез зрелых растений и проростков из их семян на крахмальном геле (это делалось для определения аллозимных фенотипов, то есть содержания аллелей некоторых сильно изменчивых ферментов, полосы которых легко обнаруживаются в электрофорезном геле по катализу специфических реакций, в результате которых образуются окрашенные продукты).
Результатом гибридизации этих осок в Европейской России является, по-видимому, то что "резкой границы между C. salina, C. recta, C. subspathacea не существует, и некоторые растения из этой группы видов не поддаются идентификации" (Егорова, 1976, 1999). В отличие от американских авторов, Егорова предполагает, что C. salina является гибридом C. recta x C. subspathacea.
Цель данной работы состояла в выделении и описании групп осок, относящихся к вышеуказанным видам, реально отличимых в полевых условиях по морфологическим признакам.
Ежегодно Беломорская экспедиция Московской Гимназии на Юго-Западе проводит исследования в окрестностях Чупинской губы, где встречаются осоки всех упомянутых видов. В июле-августе 2000 г. в окрестностях дер. Н. Пулонга проводились измерения следующих признаков осок видов C. salina, C. recta, C. subspathacea (в дальнейшем - "группа C. salina"): (1) высота максимального генеративного побега; (2-) количество мужских колосков; (3-) количество обоеполых колосков; (4-) количество женских колосков; (5) высота вегетативного побега; (6) длина кроющих чешуй; (7-) длина мешочков; (8) наличие ости на кроющих чешуях; (9) длина верхнего мужского колоска; (10) длина нижнего женского колоска; (11-) ширина носика у основания; (12-) ширина максимального листа на вегетативном побеге, измеренная в основании листа; (13) ширина нижнего прицветного листа, измеренная в основании листа; (14) наличие желобчатости нижнего прицветного листа. В дальнейшем часть признаков была отброшена (эти признаки обозначены знаком "-" в перечислении), поскольку при предварительной обработке выяснился незначительный вклад этих признаков в первые главные компоненты. В июле-августе 2002 г. в окрестностях губы Чупа и губы Кив (в том числе на островах Кемь-Лудского архипелага) проводились измерения уже по сокращенному набору признаков, измерялись виды группы C. salina, а также C. aquatilis subsp. aquatilis и C. paleacea. Всего в 2000-2002 гг. было измерено 550 особей из 22 популяций. Обработка производилась при помощи программ STATISTICA и R. Использовались различные методы многомерного анализа: кластерный и дискриминантный анализ, многомерное шкалирование и анализ главных компонент.
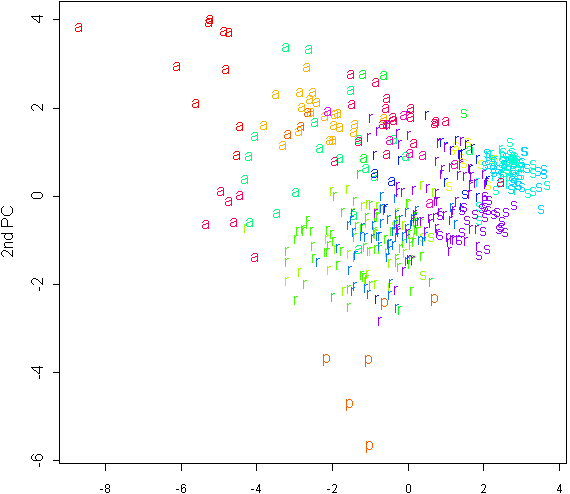
Дискриминантный анализ позволил установить, что более корректным, чем выделение трех различных видов группы C. salina, является выделение двух видов: C. recta и C. salina s.l. (включая C. subspathacea). В результате анализа главных компонент (см. рис. 1) выяснилось также, что C. salina s.l. образует очень тесную группу.
Рис. 1. График распределения измерений 2000-2002 гг. на плоскости двух первых главных компонент. Буквы соответствуют сокращенным латинским названиям видов, цвета соответствуют различным популяциям.
Для определения принадлежности видов группы C. salina был составлен следующий определительный ключ:
1. Наибольший генеративный побег длиннее 23,5 см; высота вегетативного побега вместе с наибольшим листом больше 30,5 см; верхний мужской колосок более 1,8 см в длину; нижний женский колосок более 2,4 см в длину; кроющие чешуи 5-6 мм длиной, остистые ... C. recta
-- Размеры побегов и колосков меньше приведенных в тезе, кроющие чешуи 3-4 мм длиной, остистые или без ости ... C. salina s.l. (incl. C. subspathacea)
Кроме того, по результатам обработки можно сделать следующие выводы:
В дальнейшем необходимо дальнейшее изучение взаимоотношений осок из группы C. salina с C. aquatilis и C. paleacea, поскольку наши данные пока не в позволяют выбрать одну из двух гипотез о гибридной природе этих видов. Заслуживает серьезного изучения короткостебельная форма C. aquatilis (часто выделяемая в особый подвид C. aquatilis subsp. stans), гибридизирующая с еше одним близким видом, также встречаюшимся в районе исследований - C. bigelowii (Duman, Kryszczuk, 1958). Кроме того, выраженная географическая изменчивость исследованных осок позволяет использовать их, наряду с осоками из группы C. flava, как "эволюционный маркер" при выяснении особенностей микроэволюции (особенно "эффекта основателя") на беломорских островах.
Авторы благодарят Глаголева С., Квашенко А., Волкову П., Бунтмана П., Чава В., Исаченко А., Вабищевич А., Гарушянц С., Мотылеву М., Римскую-Корсакову Н. за большую помощь в работе. Работа осуществлялась при поддержке Кандалакшского Государственного Заповедника.
Егорова Т.В. Осоки (Carex L.) России и сопредельных государств. Спб., 1999. С. 426-430.
Егорова Т.В. Осоки (Carex L.) // Флора европейской части СССР. Т. 2. Спб., 1976. С. 197.
Cayouette J., Morisset P. Chromosome studies on natural hybrids between maritime species of Carex (sections Phacocystis and Cryptocarpae) in northeastern North America, and their taxonomic implications // Can. J. Bot. 1985. Vol. 63. P. 1957-1982.
Duman M.G., Kryszczuk S.D. Introgressive hybridisation in the Carex stans-bigelowii complex // Bull. Torr. Bot. Club. 1958. Vol. 85. P. 359-362.
Haines A. Identification and taxonomy of two difficult maritime hybrids with Carex paleacea // Botanical Notes. 2000. N. 4. [Electronic resource]. Mode of access: http://www.woodlotalt.com/publications/publications.htm
Standley L. Allozyme evidence for the hybrid origin of the maritime species Carex salina and Carex recta (Cyperaceae) in Eastern North America // Syst. Bot. 1990. Vol. 15. P. 182-191.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |