1. Введение.
Кувшинка белая (Nymphaea candida L.) относится к семейству кувшинковых (Nymphaeaceae). Кувшинка — гидрофит, однако при понижении уровня воды в водоеме может жить и на суше. Водный образ жизни кувшинок, связанный с дефицитом света и кислорода, является причиной для возникновения разнолистности. У кувшинки подводные листья пленчатые, широко ланцетные, с параллельным жилкованием. Взрослые плавающие листья сердцевидные, округлые, рассеченные базальной выемкой, как правило, с длинным черешком (Дубына, 1982). Цветки одиночные, достаточно крупные, сидят на длинной цветоножке. Они обоеполые, актиноморфные, с двойным околоцветником. У цветков кувшинки наблюдается протогиния: сначала развивается гинецей, а потом — андроцей (Wiersema, 1988).
Цветки кувшинок в период цветения открываются утром и закрываются вечером. После цветения они закрываются окончательно, несколько дней плавают на поверхности воды в горизонтальном положении, а затем погружаются под воду, где и созревает плод. В некоторый момент спелый плод раскрывается, и семена, обернутые пленчатым воздухоносным присемянником, всплывают на поверхность воды. На следующий день они погружаются на дно (van der Velde, 1986).
Цветки кувшинки привлекают различных насекомых-опылителей, относящихся к отрядам двукрылые (Diptera) и перепончатокрылые (Hymenoptera) (Wiersema, 1988). Благодаря тому, что цветки закрываются вечером, насекомые остаются в цветке до его очередного открытия, поедая тычинки и обсыпаясь пыльцой, которую впоследствии переносят на другие цветки. Определение насекомых, собранных в цветках, показало, что опылителями являются береговушки (двукрылые сем. Ephydridae). Эти насекомые откладывают яйца и проводят личиночную стадию в стеблях кувшинковых.
Наблюдения за цветками кувшинки позволяют отнести их поведение к тому классу явлений который в биологической литературе получил название “биологических часов”
Выделяют три основных признака биологических часов:
При постоянных условиях и в отсутствие внешних воздействий сохраняются осцилляции с периодом около 24 часов.
Наблюдается температурная компенсация, т.е. способность поддерживать длину периода в широких пределах физиологических температур (Суини, Гастингс, 1964; Швеммле,1964; Bunning, 1931).
Часы чувствительны к переключающему стимулу, подстраивающему период к двадцатичетырехчасовому в условиях суточных изменений освещения.
Основными свойствами биологических часов, сходными практически у всех организмов, являются: фоточувствительность, непосредственное отражение субъективного времени суток величиной и направлением ответа и быстрое, в течение нескольких минут или часов, переключение (Crothwaite et al., 1995).
Среди влияющих на циркадный ритм факторов выделяют температуру, наличие питательных веществ и некоторые другие, но доминирующим сигналом для всех циркадных систем является состав и количество света (Кернер, 1895; Crothwaite et al., 1995).
В своих опытах Ф.Браун (Уинфри, 1990) показал, что листья картофеля четко реагируют на изменения атмосферного давления. Приняв во внимание, что `за фотопериодизм цветения ответственен тот же самый временной механизм, который определяет временную последовательность движения листьев' (Бюннинг, 1961), можно сделать вывод, что поведение цветков также может зависеть от атмосферного давления.
В литературе содержится мало информации о поведении цветков изучаемого нами вида. В некоторых источниках (Кернер, 1895; Калмыков, 1973; Артамонов, 1989) указано, что цветки кувшинки белой открываются в 7-8 часов, а закрываются в 17-19 часов. При этом детальная информация об характере изменения степени погруженности цветков отсутствует.
В работе были поставлены следующие задачи:
1. Изучить динамику поведения цветков кувшинки белой.
2. Выявить факторы, обуславливающие ритмику поведения цветков и интенсивность цветения кувшинки белой.
3. Сравнить ритмику поведения цветков исследуемого вида в разные годы.
4. Сопоставить опубликованные данные о биологических ритмах различных видов зеленых растений с нашими материалами по поведению цветков кувшинки белой.
2. Материалы и методы.
2.1. Общая информация.
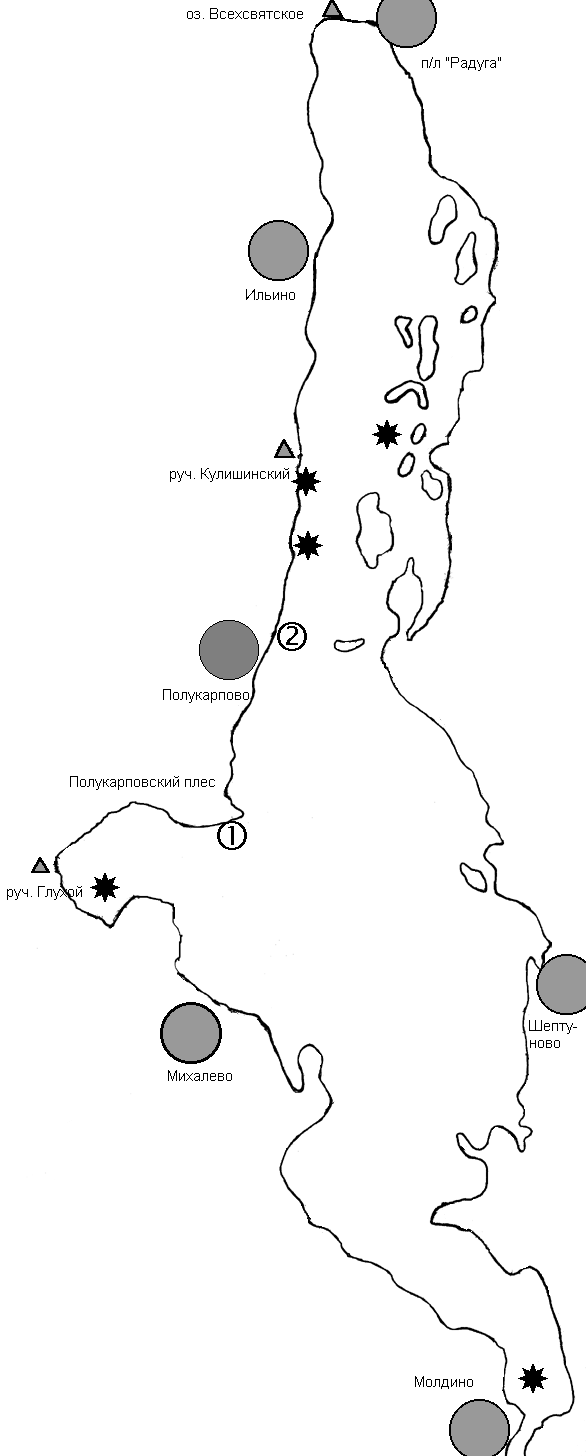
Наблюдения проводились в течение трех сезонов в сроки с 23 по 28 июня 1998 года, с 17 по 28 июня 1999 года и с 20 по 29июня 2000 года в Удомельском районе Тверской области, изучались популяции кувшинки белой на озере Молдино у деревни Полукарпово (57╟ 40' с.ш., 35╟ 10' в.д) (рис. 1).
В 1998 году 23 июня пометили пять кувшинок из одной популяции, растущих на расстоянии около 5м от берега на глубине около 130см. Четыре кувшинки пометили нитками и одну —удалением части чашелистика. Для получения большего количества данных 25 июня пометили шестую кувшинку из той же популяции удалением частей двух чашелистиков. В течение пяти дней вели наблюдения за цветками меченых кувшинок. При этом отмечали стадию развития цветков, степень их открытости и погруженности (табл. 2). Кроме того, в дни наблюдений регистрировали наличие облачности. В ходе других исследований измеряли температуру воздуха и верхнего слоя воды в центральной части озера Молдино в трехстах метрах от Полукарповского плеса (табл. 1). Всего проведено 13 наблюдений.
В 1999 году 17 июня пометили 25 кувшинок из вышеуказанной популяции. Метки представляли собой прикрепленные ниткой к цветоножке куски пенопласта размером 5х5х2см, на которые были нанесены порядковые номера растений. Подобная система мечения исключает путаницу растений и сводит к минимуму влияние меток на поведение цветков. 20 июня пометили 23 кувшинки из второй популяции, расположенной на расстоянии 1,5км от первой. 25 июня эти растения отцвели и поэтому наблюдения за ними были прекращены. В этот день пометили другие 13 кувшинок из второй популяции, за которыми наблюдали до 28 июня. Таким образом, в течение 12 дней наблюдали за поведением цветков из этих двух популяций. Кроме того, провели серию разовых наблюдений над произвольно выбранными популяциями с целью выявления межпопуляционных различий. При этом отмечали степень открытости, степень погруженности и стадию развития цветков, время суток, температуру воздуха и температуру воды, относительную влажность воздуха и облачность непосредственно на месте наблюдений. В полевом лагере (примерно в 1км от места наблюдений) цветным фитопиранометром ГОО М-80 (уловителем фотосинтетически активного солнечного излучения) в комплекте с гальванометром (измерителем постоянного тока) измеряли фотосинтетически активную радиацию (далее ФАР), по показаниям бытового барометра регистрировали атмосферное давление, а также по показаниям минимального термометра фиксировали минимальную суточную температуру (табл. 1). Всего проведено 53 наблюдения.
В 2000 году 20 июня пометили 18 кувшинок из популяции, над которой велись наблюдения в течение двух предыдущих лет. Способ мечения был избран тот же, что и в 1999 году. По мере отцветания меченых кувшинок прикрепляли метки к другим цветущим растениям этого вида. Всего в период наблюдения было помечено 56 цветков. В течение десяти дней вели наблюдения за мечеными кувшинками. В ходе каждого наблюдения отмечали степень открытости и степень погруженности цветка, один раз в сутки фиксировали стадию его развития. Кроме того, регистрировали время суток, температуру воды в окрестностях популяции, температуру воздуха, относительную влажность воздуха, атмосферное давление и облачность непосредственно на месте наблюдений. В полевом лагере измеряли фотосинтетически активную и полную солнечную радиацию (табл. 1). Измерения полной солнечной радиации проводили при помощи черно-белого пиранометра М-80 (уловителя солнечной радиации) в комплекте с гальванометром. Всего проведено 76 наблюдений.
В период с 1998 по 2000 годы наблюдения проводились с 3 до 23 часов, количество наблюдений в сутки изменялось от 4 до 14 раз. Всего проведено 142 наблюдения.
Полученные в ходе наблюдений данные обработали при помощи пакета STATISTICA for Windows (StatSoft Inc., 1995). Для проведения статистического анализа данных время наблюдений перевели в абсолютное (каждый момент времени характеризуется количеством часов, прошедших с начала наблюдений). Вычислили `средний балл степени открытости' (среднее арифметическое баллов открытости всех цветков за одно наблюдение), `средний балл степени погруженности' (среднее арифметическое баллов погруженности всех цветков за одно наблюдение) и `средний балл стадии развития' (среднее арифметическое баллов стадии развития всех цветков за одно наблюдение) для того, чтобы получить представление об общих тенденциях поведения цветков. Рассчитали коэффициенты корреляции между степенью открытости цветков, степенью погруженности цветков, временем наблюдения, температурой воды, температурой воздуха, облачностью, относительной влажностью воздуха и ФАР, а для 2000 г. и полной солнечной радиацией. Поскольку распределение значений этих характеристик было близким к нормальному, анализировались параметрические коэффициенты корреляции. К временным рядам степени открытости и погруженности, полученным в ходе наблюдений в 2000 г., был применен спектральный анализ.
Построили графики зависимости баллов `средней степени открытости' и `средней степени погруженности' от времени наблюдения с учетом стадии развития цветка (рис. 2,3).
Измерения солнечной радиации проводились при стабильном солнечном сиянии (радиация, которая остается постоянной как минимум 1 мин). Суммарную радиацию вычисляли как сумму значений прямой и рассеянной радиации. В пасмурную погоду эти значения совпадают. Пересчет значений солнечной радиации, зафиксированных по показаниям гальванометра, из условных единиц измерения в кВт/м2 проводили при помощи компьютерной программы, которая была написана канд. геогр. наук Шиловцевой О.А.
2.2. Погодные условия в период наблюдений.
Погодные условия во время полевых наблюдений были различными в разные годы. Так, в июне 1998 года погода постоянно была пасмурной, часто шел дождь, июнь 1999 года был сухим и ясным, а во время наблюдений 2000 года погода часто менялась (табл. 1).
Таблица 1. Погодные условия в период наблюдений в 1998-2000 годах (усредненные показатели).
|
t воды (град. С) |
t воздуха (град. С) |
отн. влажн. (%) |
облачн. (%) |
атм. давл. (мм. рт. столба) |
сумм. ФАР (кВт/м2) |
сумм. полная радиация (кВт/м2) |
|
|
1998г |
20,2 |
14,3 |
... |
... |
... |
... |
... |
|
1999г |
26,2 |
23,5 |
45 |
25 |
751 |
0,24 |
... |
|
2000г |
19,3 |
20,6 |
78,25 |
52,9 |
745,9 |
0,19 |
42,9 |
2.3 Характеристики состояния цветка.
Степень открытости, степень погруженности и стадию развития цветка оценивали визуально по пятибалльной шкале (табл.2, 3).
Таблица 2. Критерии определения степени открытости и степени погруженности цветка.
|
балл |
открытость |
погруженность |
|
1 |
цветок полностью закрыт |
цветок плавает на поверхности воды |
|
2 |
цветок почти закрыт |
цветок погружен на 1/3 своей высоты |
|
3 |
цветок наполовину открыт |
цветок погружен на 1/2 своей высоты |
|
4 |
цветок почти открыт |
цветок погружен на 2/3 своей высоты |
|
5 |
цветок полностью открыт |
цветок полностью погружен под воду |
Таблица 3. Критерии определения стадии развития цветка.
|
название стадии |
условное обозначение |
состояние цветка |
|
стадия бутона |
b |
цветок закрыт, чашелистики тесно прилегают друг к другу |
|
пестичная стадия |
g |
цветок недавно раскрылся, пыльники незрелые, рыльцевый диск увлажнен |
|
переходная стадия |
ga |
пыльники созревают и склоняются над рыльцевым диском |
|
тычиночная стадия |
a |
рыльцевый диск сухой, все пыльники созрели, активно продуцируется пыльца |
|
конечная стадия |
0 |
отцветание; пыльца не продуцируется, рыльцевый диск сухой |
По литературным данным продолжительность стадии бутона составляет 4-7 (1,5) суток, продолжительность стадии цветения (с начала пестичной стадии до завершения тычиночной) 4-6 (1,2) суток (van der Velde, 1986).
3. Результаты и их анализ.
3.1 Поведение цветков.
В ходе анализа индивидуальной динамики изменения степени открытости и степени погруженности цветков кувшинки белой в период наблюдений с учетом их стадии развития получили следующие результаты.
На протяжении развития цветка его поведение меняется (рис.2,3).
3.1.1. Поведение цветка на стадии бутона.
По данным 2000 года на стадии бутона большинство цветков было закрыто на всем протяжении суток, что следует из определения этой стадии развития. На стадии бутона в большинстве случаев можно наблюдать довольно четкую суточную динамику изменения степени погруженности. Цветок наполовину погружен в воду в период приблизительно с 8 до 17 часов и полностью погружен под воду в остальные часы. Следует отметить, что по мере развития бутона уменьшается степень его погруженности.
3.1.2. Поведение цветка на стадиях цветения.
По результатам наблюдений 1999 и 2000 годов выявили четко выраженную суточную динамику изменения степени открытости и степени погруженности цветков во время их цветения, сохраняющуюся на всех его стадиях (от пестичной до тычиночной). Поведение цветков в разные годы различается.
По данным 1999 года цветки открываются и поднимаются на поверхность в 6-8 часов, закрываются и погружаются в 16-18 часов. В 2000 году для большинства цветков отмечена следующая динамика изменения степени погруженности и степени открытости цветков в течение суток. Цветки начинают открываться и подниматься на поверхность воды в 7-9 часов. Степень открытости достигает своего максимального значения в 12-13 часов, после чего цветки начинают закрываться. Цветки закрываются полностью и погружаются под воду в 17-19 часов. При этом цветки поднимаются на поверхность воды приблизительно в 8 часов и начинают погружаться лишь приблизительно в 15 часов, когда они уже наполовину закрыты. Во время цветения цветки, как правило, не погружаются более, чем на половину своей высоты.
3.1.3. Поведение цветка на конечной стадии.
Наблюдения 1998-2000 годов показали, что при отцветании динамика изменения степени открытости и степени погруженности цветков становится нечеткой, а впоследствии исчезает. При этом, как правило, цветок практически полностью погружается под воду, оставаясь наполовину открытым.
Надо отметить, что поведение не всех цветков совпадало с вышеописанным. Иногда наблюдали несколько максимумов степени открытости или степени погруженности цветков в течение суток или другие нарушения характерной для большинства цветков динамики изменения степени открытости и степени погруженности. Таким образом, анализ зависимости `средней степени открытости' и `средней степени погруженности' цветков от времени наблюдения с учетом стадии их развития позволяет выявить исключительно общие тенденции поведения цветков в популяции в целом.
3.1.4. Зависимость степени открытости цветков от степени их погруженности.
Результаты анализа данных 1998 и 1999 годов показали, что степень открытости цветков обратно пропорциональна степени их погруженности, то есть открытый цветок, как правило, плавает на поверхности воды и, наоборот, закрытый цветок в большинстве случаев погружен. В 2000 году наблюдалась иная картина — на стадии цветения зависимость этих величин нелинейная, а на стадии бутона и стадии отцветания она отсутствует. Спектральный анализ временных рядов степени открытости и степени погруженности показал, что степень открытости изменяется с периодичностью в сутки без четко выраженного тренда, а для степени погруженности наряду с суточной периодичностью выявлен нелинейный тренд. Суть этого тренда состоит в том, что минимальная степень погруженности цветков достигается на стадии цветения, переходной от пестичной к тычиночной.
3.2. Факторы, обуславливающие поведение цветков.
В ходе серии разовых наблюдений над различными популяциями кувшинки белой озера Молдино в 1999 году не удалось выявить межпопуляционных различий в характере динамики изменения открытости и погруженности цветков.
Из описания поведения цветка на стадиях бутона, цветения и отцветания видно, что динамика изменения открытости и погруженности цветка на этих стадиях различна. Это значит, что стадия развития цветка определяет динамику изменения его открытости и погруженности.
При анализе параметрических коэффициентов корреляции между `средней степенью открытости', `средней степенью погруженности' цветков и погодными условиями установили следующую взаимосвязь этих величин. Степень открытости цветков прямо пропорциональна значениям солнечной радиации (как фотосинтетически активной, так и полной), температуры воды и температуры воздуха, в то время как степень погруженности цветков обратно пропорциональна значениям этих природных факторов. Изменение `средней степени открытости' цветков в течение суток сходно с изменением значений температуры воды и полной солнечной радиации по данным 2000 года. Отсюда можно сделать вывод, что регуляция амплитуды изменения открытости и погруженности цветков в течение суток в период цветения осуществляется природными факторами, имеющими суточный ход, такими как солнечная радиация, температура воды и температура воздуха.
По данным 1999 года в пасмурную погоду, когда атмосферное давление ниже нормального, продолжительность `дня' кувшинки (время, когда цветок открыт и плавает на поверхности) меньше, чем в ясную погоду (8 и 9,5 часов соответственно). В ясную погоду, при одинаковой ФАР, `день' кувшинки короче при более низком давлении (при атмосферном давлении 742мм ртутного столба, продолжительность `дня' составляет 7 часов; при атмосферном давлении 756мм ртутного столба, продолжительность `дня' -- 11 часов). Из этого следует, что продолжительность `дня' кувшинки по данным 1999года прямо пропорциональна атмосферному давлению. В 2000 году в период цветения атмосферное давление изменялось незначительно, поэтому мы не имеем возможности анализировать зависимость продолжительности `дня' кувшинки от атмосферного давления.
У многих цветков на третьи сутки наблюдений в 2000 году на стадии цветения наблюдали нетипичную ритмику изменения степени открытости цветков. Цветки открывались полностью уже приблизительно к 9 часам и оставались открытыми примерно до 15часов. В этот день зарегистрировали наибольшую облачность (в среднем за день 95%, при средней облачности за все время наблюдений 53%). При этом цветки были полностью открытыми дольше, чем в другие дни. Отсюда следует, что продолжительность `дня' кувшинки по данным 2000 года прямо пропорциональна проценту облачности. В 1999 году облачность в период наблюдений была перистой, поэтому изменение процента облачности слабо влияло на освещенность и не воспринималось растениями. Этим может объясняться отсутствие влияния процента облачности на длину `дня' кувшинки в 1999 году.
3.3 Факторы, влияющие на интенсивность цветения.
Сопоставляя данные разных лет, мы можем отметить следующие факты.
В 1998 году при холодной и пасмурной погоде (табл. 1) цветение проходило в среднем за 6-10 суток. В 1999 году при теплой и ясной погоде цветение проходило в среднем за 3-4 суток. В частности, продолжительность пестичной стадии составляет 1-2 суток, а продолжительность переходной стадии равна продолжительности тычиночной стадии и составляет в среднем одни сутки. В 2000 году при переменчивой погоде цветение проходило в среднем за 5-6 суток (рис. 2,3). При этом продолжительность пестичной стадии составляет 2-3 суток, продолжительность переходной стадии — 2 суток, а продолжительность тычиночной стадии — 1-2 суток.
Это значит, что интенсивность цветения растений зависит от температуры воздуха и температуры воды и, возможно, от других природных факторов, таких как атмосферное давление, солнечная радиация и т.п.
4. Обсуждение результатов.
Существует несколько объяснений причин изменения степени открытости и степени погруженности цветков кувшинки белой в течение суток. По одной из самых распространенных гипотез степень погруженности цветков кувшинки белой обуславливается длиной цветоноса растения. Длина цветоноса достигает 1,5 м, поэтому даже незначительные изменения в размере (объеме) отдельных клеток приводят к заметным изменениям длины цветоноса и, как следствие, к изменению степени погруженности цветка. Изменение размера клеток чашелистиков и/или лепестков влечет за собой изменение степени открытости цветка. Известно, что объем клеток прямо пропорционален их тургору. Осморегуляция в хлорофиллоносных клетках — результат трех метаболических процессов: ионного транспорта (иона калия и сопровождающих анионов), образования сахаров при фотосинтезе и деградации крахмала с образованием сахарозы (Tallman, Zeiger, 1988). Таким образом, из факторов, влияющих на состояние клеток, а, следовательно, на поведение цветков, основным является интенсивность метаболических процессов в клетке и в том числе — интенсивность фотосинтеза.
По нашим данным, динамика изменения длины цветоноса определяется стадией развития цветка, а степень этого изменения в течение суток на стадии цветения регулируется значениями солнечной радиации и/или другими природными факторами, имеющими суточный ход и влияющими на интенсивность процессов метаболизма в клетках цветка и цветоноса. Эта гипотеза находит подтверждение в работах Кернера (1895), Уинфри (1990) и Уилкинса (1964).
Скорость изменения длины цветоноса обусловлена атмосферным давлением, что можно объяснить следующим образом. Чем выше атмосферное давление, тем медленней изменяется тургор, размер отдельных клеток, состояние цветка и цветоноса в целом, и, следовательно, степень погруженности и степень открытости цветка, тем продолжительней `день' кувшинки. Этим подтверждается предположение Bunning (1931) и выводы из опытов Брауна (Уинфри, 1990). Менее активное по сравнению с наблюдаемым в другие дни изменение степени открытости цветков при сильной облачности в 2000 году может быть также объяснено медленным изменением тургора, вызванным низкой интенсивностью фотосинтеза в условиях недостаточной освещенности.
Обратно пропорциональное изменению степени погруженности изменение степени открытости цветка в период цветения, является способом защитить пыльцу от намокания, тем самым сохраняя ее пригодной для опыления. При отцветании, когда опыление уже произошло, ритмика исчезает, и вода свободно проникает в цветок, что согласуется с предположением о связи суточной ритмики открытости цветков с опылением. Кроме того, закрывание цветка на ночь на стадии цветения позволяет удерживать внутри него опылителей для повышения эффективности опыления.
В ходе разовых наблюдений над цветками Nymphaea tetragona Georgi в условиях полярного дня (Карелия, окрестности оз. Белое, 2000 год) обнаружили отклонения от описанного в настоящей работе поведения цветков Nymphaea candida L. В частности, неоднократно наблюдали открытые ночью цветки. В дальнейшем предполагается изучить межвидовые различия в поведении цветков растений рода Nymphaea L. и влияние на поведение цветков длины светового дня.
Наличие цветков, изменяющих степень открытости и степень погруженности в период наблюдений отлично от большинства цветков, позволяет предположить существование не учитывавшихся нами факторов, которые влияют на поведение некоторых цветков. В отдельно взятых случаях возможна также неточная визуальная оценка состояния цветков.
Скорость смены стадий цветения, то есть продолжительность цветения, зависит от скорости созревания пыльников. В основе этого процесса лежат, по-видимому, биохимические реакции, скорость которых прямо пропорциональна температуре (Швеммле,1964; Bunning, 1931) и, возможно, зависит от некоторых других природных факторов.
Таким образом наша работа позволяет сделать следующие выводы:
1. Поведение цветков кувшинки белой определяется стадией их развития.
2. Регуляция амплитуды изменения открытости и погруженности цветков кувшинки белой в течение суток на стадии цветения осуществляется природными факторами, имеющими суточный ход.
3. Продолжительность `дня' кувшинки белой зависит от атмосферного давления и облачности.
4. Интенсивность цветения кувшинки белой зависит от температуры воздуха и температуры воды.
5. Благодарности.
Работа велась в рамках летних биологических практик гимназии на Юго-Западе (╧ 1543). Хочется выразить признательность старшему научному сотруднику метеорологической обсерватории МГУ канд. геогр. наук О. А. Шиловцевой за предоставленное метеорологическое оборудование и консультации по его использованию и обработке данных. Полезную информацию о методах статистической обработки данных предоставил канд. биол. наук Пузаченко А.Ю. В проведении полевых наблюдений принимали деятельное участие Абрамова Л., Григорьева Т., Кольчугина Е., Леоненко Е., Молчанов И., Селиванова А., Снеткова Н., Шашкова А.
Литература.
Артамонов В.И. Зеленые оракулы. М., 1989. 115 с.
Бюннинг Э. Ритмы физиологических процессов. М., 1961. С. 65.
Дубына Д.В. Кувшинковые Украины. Киев, 1982. 198 с
Калмыков К.Ф. Биологические часы // Детская Энциклопедия. Т. 4. М., 1973. С. 65-67.
Кернер, фон Марилаун А. Жизнь растений. Т. 2. Спб, 1895. С. 196-206.
Кувшинка; Кувшинковые // Биологический энциклопедический словарь, М., 1986. С. 301-302.
Снигиревская Н. С. Кувшинковые // Жизнь растений. Т.5, Ч.2. М., Просвещение, 1981. С. 171-175.
Суини Б., Гастингс Дж. Влияние температуры на суточные ритмы // Биологические часы. М., 1964. С. 152-155.
Уилкинс М. Влияние света на ритмы растений // Биологические часы, М., 1964. С. 154-155.
Уинфри А.Т. Время по биологическим часам. М., 1990. 315 с.
Швеммле Б. Термопериодические эффекты и цветение растений // Биологические часы. М., 1964. С. 27-35.
Bunning E. Untersuchungen uber die automen tagesperiodischen Bewegungen der Primarblatter von Phaseolus multiflorus // Jahrb. Wiss. Bot. 1931. Bd. 75. S. 439-480.
Crothwaite S.K., Loros J.J., Dunlap J.C. Light-induced resetting of a circadian clock is mediated by a rapid increase in frequency transcript // Cell. 1995. Vol. 81. P. 1003-1012.
Tallman G., Zeiger E. Light quality and osmoregulation in Vicia guard cell // Plant Physiol. 1988. Vol. 88. P. 887-895.
Velde, van der G. Developmental stages in the floral biology s. l. of dutch Nymphaeaceae (Nymphaea alba L., Nymphaea candida Presl, Numphar lutea (L.) Sm.) // Acta Bot. Neerl. 1986. Vol. 35, N 2. P. 111-113.
Wiersema J.H. Reproductive biology of Nymphaea (Nymphaeaceae) // Annals of the Missouri Botanical Garden. 1988. Vol. 75, N 3. P. 795-804.
Подписи к иллюстрациям.
Рисунок 1. Карта района исследований.
Условные обозначения: 1 -- модельная популяция (`первая') кувшинки белой, за которой велись наблюдения в 1998-2000 годах
2 -- модельная популяция (`вторая') кувшинки белой, за которой велись наблюдения в 1999 году
Звездочками обозначены обнаруженные в ходе картирования популяции кувшинки белой.
Рисунок 2. Изменение степени открытости цветков кувшинки белой в период наблюдений по данным 2000 года.
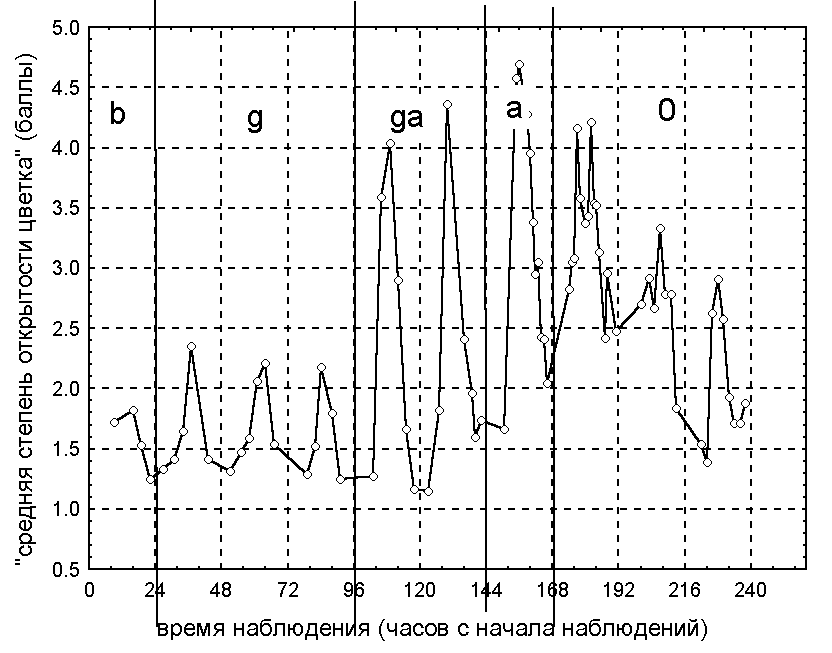
условные обозначения:
стадии цветения:
b - стадия бутона;
g - пестичная;
ga - переходная;
a - тычиночная;
0 - конечная.
Рисунок 3. Изменение степени погруженности цветков кувшинки белой в период наблюдений по данным 2000 года.
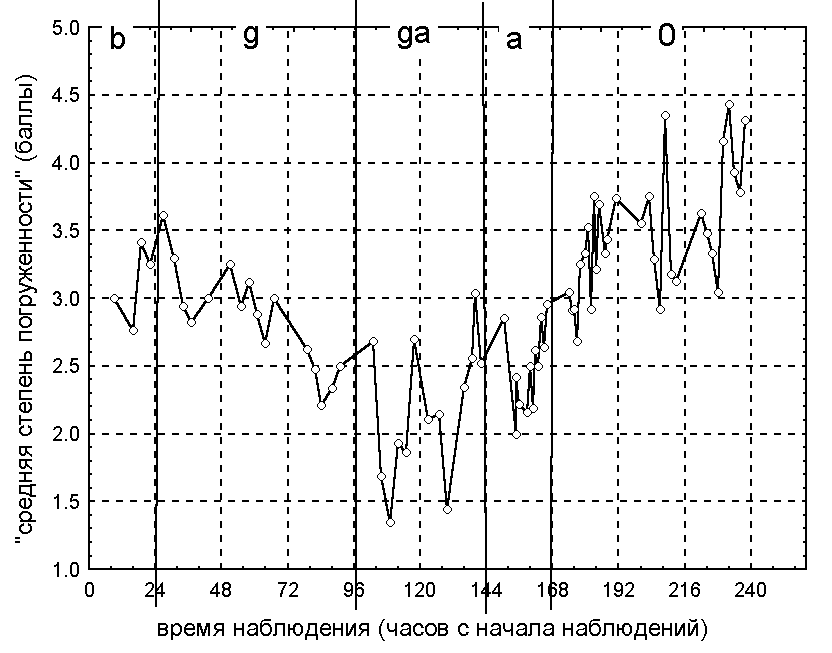
Условные обозначения: см. условные обозначения к рис. 2.