[Презентация (PDF-файл] [2001 год] [2002 год]
Сложность систематики осок, относящихся к секции Phacocystis связана с межвидовой гибридизацией, поэтому отдельные растения с трудом поддаются определению [1, 2, 6, 7]. Carex salina, по мнению многих авторов, является видом гибридного происхождения (стабилизированным гибридом) от C. subspathacea × C. paleacea [7, 10, 5]. Эта гипотеза основывается на таких признаках, как хромосомные числа, изоморфные аллельные ферменты (allozyme), промежуточная морфология, характер распространения. Последние авторы предполагают также гибридную природу C. recta (C. paleacea × C. aquatilis). Существует иное мнение, что C. salina - продукт от скрещивания C. recta × C. subspathacea [1].
Хотя большинство гибридов, представленных в секции Phacocystis и в других секциях, скорее всего, являются спорадическими гибридами первого поколения, которые либо стерильны, либо имеют пониженную фертильность, некоторые гибриды из секции Phacocystis становятся стабилизированными и широко распространенными. Они морфологически отличимы от гибридов первого поколения и дают жизнеспособные семена [7, 10]. Предполагаемые природные гибриды часто описываются на основе промежуточной морфологии. Репродуктивная изоляция среди видов осок внутри одной секции или, в некоторых случаях, среди родственных секций, вызывается, по мнению некоторых исследователей, скорее экологическими и фенологическими различиями, чем генными или хромосомными [17]. Стабилизированные гибриды сохраняют способность размножаться вегетативно и занимают различные экологические ниши. Подобная гибридизация чаще происходит в северных районах, в частности, из-за нарушения фенологической изоляции [17].
Цель данной работы состоит в выделении и описании подгрупп осок группы C. salina, реально отличимых "в поле" по указанным в определителях морфологическим признакам. В работе приводится также попытка продемонстрировать возможность использования в качестве маркеров микроэволюции (в условиях изоляции на разных островах) признаков этих и некоторых других видов растений, отличающихся большой изменчивостью.
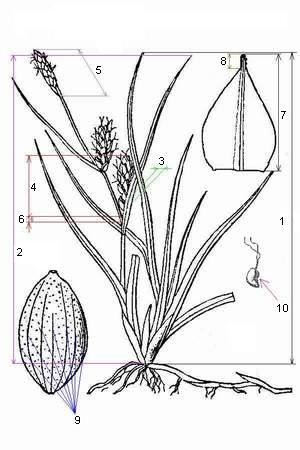
В 2000 и 2002 годах на побережье Белого моря (губа Чупа, губа Кив, Кемь-Лудские о-ва и о-ва Керетского архипелага) мы проводили измерения морфологических признаков (рис. 2) осок пяти указанных видов. Экземпляров C. paleacea было найдено крайне мало, всего 7, к тому же все они отчетливо отличались от всех остальных видов [1], поэтому эти данные были исключены из рассмотрения. В 2003 г. собирались образцы из популяций этих осок, а также, чтобы сравнить характер изменчивости признаков, собирались образцы популяций С. serotina s.l., Achillea millefolium и Euphrasia spp., растущих на островах Кив-губы Белого моря (эти виды также известны своей вариабильностью [4, 6, 8, 11, 16, 18]). Был исследован 31 остров, причем на всех островах присутствовали популяции Euphrasia, только на 19-и островах имелись популяции Achillea и всего на 5-и островах были встречены осоки секции Phacocystis (C. aquatilis), а на 4-х островах были найдены популяции C. serotina s.l. Всего было измерено 671 растение осок секции Phacocystis из 28 популяций по следующим 11 диагностическим признакам:
Обработка проводилась в пакете STATISTICA 5.5 [14], а также в статистической среде R 1.8.1 [9]. Применялся анализ главных компонент, кластерный, дискриминантный, корреляционный анализ и таксономический анализ Смирнова, а также использовались описательные статистики [3, 13].
Многомерные методы не поддерживают отсутствующие данные, которые при анализе пропускались. Наиболее часто отсутствовал вегетативный побег.
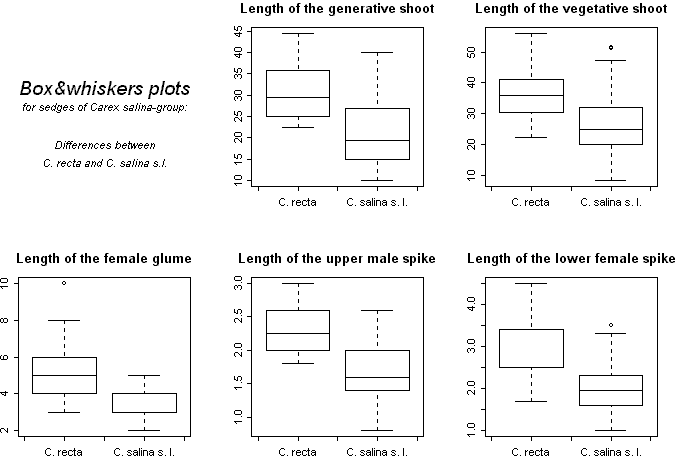
Сначала был проведен предварительный анализ. В ходе него измеренные образцы 2000 года (75 особей) были разбиты на виды в соответствии с ключом [1]. При применении дискриминантного анализа к этим данным получена классификация, которая существенно отличается от исходной, учитывая малый объем выборки (табл. 1). Была произведена попытка "образовать" из трех плохо различимых видов (C. salina, C. recta, C. subspathacea) различные группы — C. recta и C. "salina-subspathacea" ("вид", к которому отнесены предполагаемые представители C. salina, и C. subspathacea) или C. "salina-recta" и C. subspathacea. Затем было проведено сравнение процентов корректности классификации, полученных при помощи дискриминантного анализа. Наиболее корректным является выделение — C. recta, C. "salina-subspathacea", поскольку в этом случае наблюдается наибольший общий процент корректности — 90,54 (табл. 2). Для разных видов составлены диаграммы размаха (box & whiskers plot) по непрерывным признакам (рис. 1).
| Группы | Процент корректности | "recta" | "salina" | "subspathacea" |
| p = 0,32432 | p = 0,33784 | p = 0,33784 | ||
| "recta" | 87,50 | 21 | 2 | 1 |
| "salina" | 72,00 | 3 | 18 | 4 |
| "subspathacea" | 92,00 | 0 | 2 | 23 |
| Для всех | 83,78 | 24 | 22 | 28 |
| Группы | Процент корректности | "recta" | "salina-subspathacea" |
| p = 0,32432 | p = 0,67568 | ||
| "recta" | 83,33 | 20 | 4 |
| "salina-subspathacea" | 94,00 | 3 | 47 |
| Для всех | 90,54 | 23 | 51 |
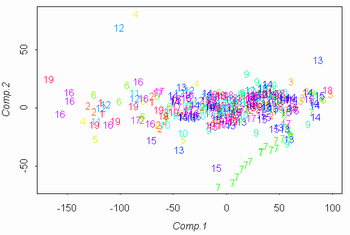
Затем был проведен анализ данных 2002 года. В ходе него данные были обработаны при помощи анализа главных компонент (рис. 5а). В построенном на осях главных компонент графике обозначения соотвествуют видам осок. При этом наибольшие нагрузки на главные компоненты несут такие признаки, как длина генеративного и вегетативного побега. Однако эти признаки могут сильно коррелировать со внешними условиями или просто отличаться большой внутрипопуляционной изменчивостью. Поэтому при исключении из рассмотрения этих признаков получается другой график (рис. 5б).
Рисунок 1. Диаграмма размаха по непрерывным признакам для C. recta и C. salina s. l. |
Рисунок 2. Измеряемые признаки осок (обозначения признаков см. выше). |
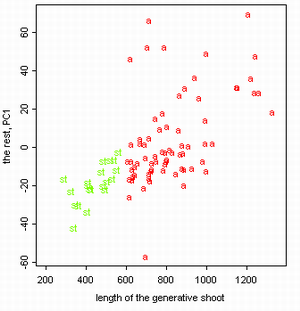 | 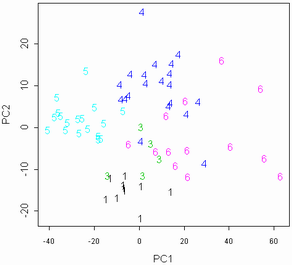 |
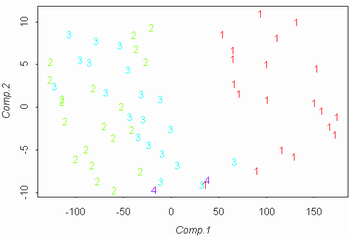
Рисунок 3. Разделение C. aquatilis subsp. aquatilis ("a") и C. aquatilis subsp. stans ("st"). |
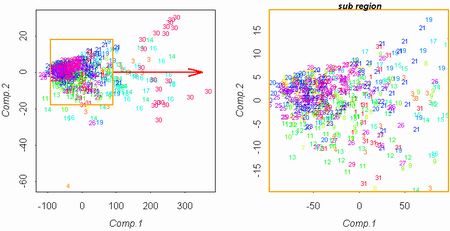
Рисунок 4. Разделение "маркеров" на популяции (цифры соответствуют номерам популяций). |
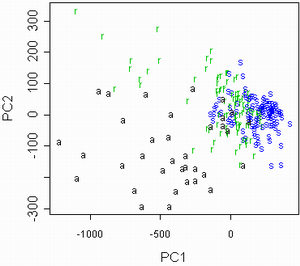 | 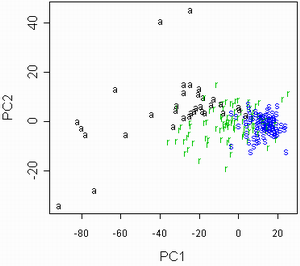 |
Рисунок 5. Разделение данных 2002 года на виды ("a" = C. aquatilis; "r" = C. recta; "s" = C. salina s.l.)
| |
1. Наибольший генеративный побег длиннее 23,5 см; высота вегетативного побега вместе с наибольшим листом превышает 30,5 см; верхний мужской колосок более 1,8 см дл.; нижний женский колосок более 2,4 см дл.; кроющие чешуи 5-6 мм дл., остистые ---- C. recta
- Размеры побегов и колосков меньше приведенных в тезе, кроющие чешуи 3-4 мм дл., остистые или без ости ---- C. salina s.l. (incl. C. subspathacea)
Результаты обработки показали, что разные популяции отличаются друг от друга отчетливее, чем виды, причем существенно лучше популяции различаются, если убрать из рассмотрения такие признаки, как длины побегов (рис. 4, 5). Благодаря настолько сильно выраженным межпопуляционным отличиям, осоки секции Phacocystis могут выступать в качестве "маркеров микроэволюции" (этот термин в данной работе обозначает изучаемые изолированные популяции — ничего общего с молекулярными маркерами не имеется в виду) в условиях изоляции. Такая изоляция имеет место среди популяций осок, произрастающих на разных островах.
Рисунок 6. Разделение Achillea на популяции
|
Рисунок 7. Разделение Euphrasia на популяции (цифры соответствуют номерам популяций). |
Как видно, и образцы из популяций Achillea, и образцы из популяций Euphrasia сильно смешиваются — разные популяции почти неразличимы. Это особенно показательно на примере тех популяций, которые произрастают там же, где соответствующие популяции осок — осоки разделяются лучше. Скорее всего, это связано с тем, что широкая норма реакции (дисперсия признаков) перекрывает имеющиеся у этих групп межпопуляциюнные различия. Зато отдельные популяции каждой из групп осок различимы; возможно это объясняется небольшой величиной выборки, но не исключено, что межпопуляционные различия действительно выше у осок, чем у других изменчивых видов.
Рисунок 8. Разделение C. serotina на популяции
|
Работа проводилась при содействии Кандалакшского государственного заповедника.
[1] Егорова Т.В. Осоки (Carex L.) России и сопредельных государств. Спб., 1999. С. 426-430.
[2] Егорова Т.В. Флора европейской части СССР. Спб., 1976. С. 197.
[3] Шмидт В.М. Математические методы в ботанике. Спб., 1984. 288 с.
[4] Blackstock N., Ashton P.A. A re-assesment of the putitave Carex flava agg. (Cyperaceae) hybrids at Malham Tarn (v.c. 64): A morphometric analysis // Watsonia. 2001. Vol. 23. P. 505-516.
[5] Cayouette J., Morisset P. Chromosome studies on natural hybrids between maritime species of Carex (sections Phacocystis and Cryptocarpae) in northeastern North America, and their taxonomic implications // Can. Journ. Bot. 1985. Vol. 63. P. 1957-1982.
[6] Duman M.G., Kryszczuk S.D. Introgressive hybridization in the Carex stans-bigelowii complex // Bull. Torr. Bot. Club. 1958. Vol. 85. P. 359-362.
[7] Haines A. Identification and taxonomy of two difficult maritime hybrids with Carex paleacea // Botanical Notes. 2000. N. 4. [Electronic resource]. Mode of access: http://www.woodlotalt.com/publications/publications.htm.
[8] Pyk J., Toinoven H. Taxonomy of the Carex flava complex (Cyperaceae) in Finland // Nord. J. Bot. 1994. Vol. 14. P. 173-191.
[9] R Development Core Team. R: A language and environment for statistical computing // R Foundation for Statistical Computing, Vienna, Austria [Electronic resource]. Mode of access: http://www.R-project.org.
[10] Standley L. Allozyme evidence for the hybrid origin of the maritime species Carex salina and Carex recta (Cyperaceae) in Eastern North America // Syst. Bot. 1990. Vol. 15. P. 182-191.
[11] Saukel J., Langer R. Die Achillea millefolium-Gruppe (Asteraceae) in Mitteleuropa // Phyton. 1992. Vol. 31. P. 185-207.
[12] Schmid B. Notes on the nomenclature and taxonomy of the Carex flava group in Europe // Watsonia. 1983 Vol.14. P. 309-319.
[13] S-PLUS 2000 Guide to Statistics // Data Analysis Products Division MathSoft, Inc. 1999. Vol. 2. Seattle, Washington.
[14] STATISTICA for Windows [Computer program manual] // StatSoft, Inc. 1999. Tulsa, OK.
[15] StatSoft, Inc. (2001). Электронный учебник по статистике. Москва, StatSoft. WEB: http://www.statsoft.ru/home/textbook/default.htm.
[16] Swann E.L. Norfolk Euphrasiae // Watsonia. 1983. Vol. 9. P. 345-356.
[17] Whitkus R. Experimental hybridizations among chromosome races of Carex pachystachya and related species C. macloviana and C. preslii (Cyperaceae) // Syst. Bot. 1988. Vol. 13. P. 146-153.
[18] Yeo P.F. A taxonomic revision of Euphrasia in Europe // Bot. Journ. Linn. Soc. 1978. Vol. 77. P. 223-334.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |